Difference between revisions of "Salix tracyi"
Univ. Calif. Publ. Bot. 17: 403, plates 69, 70. 1934.
FNA>Volume Importer |
FNA>Volume Importer |
||
| Line 24: | Line 24: | ||
|elevation=90-500 m | |elevation=90-500 m | ||
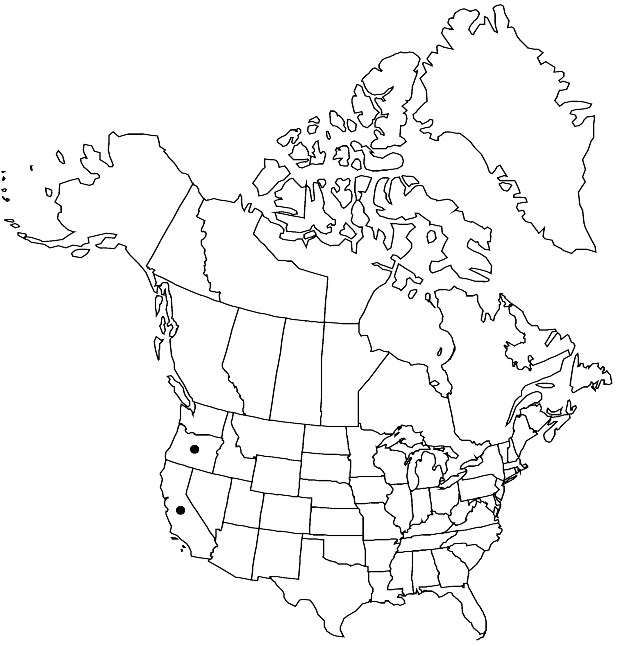
|distribution=Calif.;Oreg. | |distribution=Calif.;Oreg. | ||
| − | |discussion=<p>Ball described Salix tracyi as part of a group of “more or less localized endemic willows of northern California and southern Oregon.” A. K. Skvortsov (1971), followed by G. W. Argus (1993), thought that this species was a stunted form, perhaps shade-growth, of S. lasiolepis. R. D. Dorn (2000) studied the taxon in the field and considered it to be a distinct species. He noted that it could “usually” be separated from S. lasiolepis by its shorter styles, longer stipes, less hairy floral bracts, shorter catkins, thinner, less hairy leaves, and reddish juvenile leaves. He also noted differences in flowering times but they overlap completely. The differences are subtle and, although not diagnostic, should be studied further. Salix tracyi is treated here as a species because of its relatively large, localized range and interesting biological and morphological characteristics. Local botanists making collections from tagged individuals could provide useful information on habitat, elevation, flowering time, and morphology. Experimental study of this and the other serpentine endemic Salix in the region (see also 110. S. breweri and 111. S. delnortensis) could provide insights into their evolution and their relationship with S. lasiolepis.</p><!-- | + | |discussion=<p>Ball described <i>Salix tracyi</i> as part of a group of “more or less localized endemic willows of northern California and southern Oregon.” A. K. Skvortsov (1971), followed by G. W. Argus (1993), thought that this species was a stunted form, perhaps shade-growth, of <i>S. lasiolepis</i>. R. D. Dorn (2000) studied the taxon in the field and considered it to be a distinct species. He noted that it could “usually” be separated from <i>S. lasiolepis</i> by its shorter styles, longer stipes, less hairy floral bracts, shorter catkins, thinner, less hairy leaves, and reddish juvenile leaves. He also noted differences in flowering times but they overlap completely. The differences are subtle and, although not diagnostic, should be studied further. <i>Salix tracyi</i> is treated here as a species because of its relatively large, localized range and interesting biological and morphological characteristics. Local botanists making collections from tagged individuals could provide useful information on habitat, elevation, flowering time, and morphology. Experimental study of this and the other serpentine endemic <i>Salix</i> in the region (see also 110. <i>S. breweri</i> and 111. <i>S. delnortensis</i>) could provide insights into their evolution and their relationship with <i>S. lasiolepis</i>.</p><!-- |
| − | --><p>A numerical taxonomic study by T. J. Crovello (1968) found that Salix tracyi clustered with members of sect. Cordatae, but both G. W. Argus (1997) and R. D. Dorn (2000) placed it in sect. Mexicanae with S. lasiolepis.</p> | + | --><p>A numerical taxonomic study by T. J. Crovello (1968) found that <i>Salix tracyi</i> clustered with members of sect. Cordatae, but both G. W. Argus (1997) and R. D. Dorn (2000) placed it in sect. Mexicanae with <i>S. lasiolepis</i>.</p> |
|tables= | |tables= | ||
|references= | |references= | ||
| Line 49: | Line 49: | ||
|publication year=1934 | |publication year=1934 | ||
|special status= | |special status= | ||
| − | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/ | + | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/8f726806613d60c220dc4493de13607dd3150896/coarse_grained_fna_xml/V7/V7_181.xml |
|genus=Salix | |genus=Salix | ||
|subgenus=Salix subg. Vetrix | |subgenus=Salix subg. Vetrix | ||
Revision as of 17:55, 18 September 2019
Shrubs, 1–6 m. Stems: branches gray-brown to red-brown, strongly to weakly glaucous, glabrous; branchlets yellow-brown to red-brown, (weakly glaucous), glabrous or sparsely to moderately densely velvety or tomentose. Leaves: stipules absent, rudimentary, or foliaceous on early ones, foliaceous on late ones, (2.5–9 mm), apex rounded or convex; petiole convex to flat, or shallowly grooved adaxially, 5–11 mm, tomentose or velvety adaxially; largest medial blade lorate, oblanceolate, or elliptic, 55–96 × 15–34 mm, 2–3.7 times as long as wide, base convex, margins slightly revolute, entire, sinuate or serrulate, apex acuminate to acute, abaxial surface glaucous, sparsely pubescent to moderately densely tomentose or glabrous, hairs (white, sometimes also ferruginous), straight or curved, adaxial slightly glossy, sparsely tomentose to glabrescent; proximal blade margins entire or serrulate; juvenile blade reddish, very densely tomentose or puberulent to glabrescent abaxially, hairs white, sometimes also ferruginous. Catkins flowering as or just before leaves emerge; staminate stout, 17–30 × 7–9 mm, flowering branchlet 1.5–5 mm; pistillate very densely flowered, stout to slender, 17–42 × 6–11 mm, flowering branchlet 1.5–3 mm; floral bract brown or bicolor, 0.8–1.6 mm, apex rounded to truncate, abaxially hairy throughout or proximally, hairs wavy or straight. Staminate flowers: adaxial nectary narrowly oblong, oblong, or ovate, 0.3–0.8 mm; filaments connate less to more than 1/2 their lengths; anthers purple turning yellow, ellipsoid, shortly cylindrical, or globose, 0.36–0.44 mm. Pistillate flowers: adaxial nectary oblong, or narrowly oblong to flask-shaped, 0.2–0.6 mm; stipe 1–1.7(–2.4) mm; ovary pyriform, beak sometimes slightly bulged below styles; ovules 12 per ovary; styles 0.1–0.6 mm; stigmas flat, abaxially non-papillate with rounded or pointed tip, 0.1–0.3 mm. Capsules 2.4–3.6 mm.
Phenology: Flowering early Apr-early May.
Habitat: Shores and floodplains, sandy, gravelly, or rocky substrates, often serpentine
Elevation: 90-500 m
Discussion
Ball described Salix tracyi as part of a group of “more or less localized endemic willows of northern California and southern Oregon.” A. K. Skvortsov (1971), followed by G. W. Argus (1993), thought that this species was a stunted form, perhaps shade-growth, of S. lasiolepis. R. D. Dorn (2000) studied the taxon in the field and considered it to be a distinct species. He noted that it could “usually” be separated from S. lasiolepis by its shorter styles, longer stipes, less hairy floral bracts, shorter catkins, thinner, less hairy leaves, and reddish juvenile leaves. He also noted differences in flowering times but they overlap completely. The differences are subtle and, although not diagnostic, should be studied further. Salix tracyi is treated here as a species because of its relatively large, localized range and interesting biological and morphological characteristics. Local botanists making collections from tagged individuals could provide useful information on habitat, elevation, flowering time, and morphology. Experimental study of this and the other serpentine endemic Salix in the region (see also 110. S. breweri and 111. S. delnortensis) could provide insights into their evolution and their relationship with S. lasiolepis.
A numerical taxonomic study by T. J. Crovello (1968) found that Salix tracyi clustered with members of sect. Cordatae, but both G. W. Argus (1997) and R. D. Dorn (2000) placed it in sect. Mexicanae with S. lasiolepis.
Selected References
None.