Selaginella rupestris
Flora 21: 182. 1838.

Plants on rock or terrestrial, forming long or spreading mats or rarely cushionlike mats. Stems radially symmetric, long to moderately short-creeping to decumbent, not readily fragmenting, irregularly forked, without budlike arrested branches, tips straight; main stem indeterminate, lateral branches conspicuously or inconspicuously determinate, sometimes ascending, 1–3-forked. Rhizophores borne on upperside of stems, throughout stem length, 0.25–0.45 mm diam. Leaves monomorphic, in alternate pseudowhorls of 6 (on main stem) to 4 (on lateral branches), tightly appressed, ascending, green, occasionally reddish, linear or linear-lanceolate, 2.5–4(–4.5) X 0.45–0.6 mm; abaxial ridges well defined; base cuneate and decurrent on underside to rounded and adnate on upperside, pubescent or glabrous; margins long-ciliate, cilia transparent, spreading, (0.05–)0.07–0.17 mm; apex slightly keeled, mostly attenuate; bristle white, whitish, or transparent, puberulent, 0.45–1(–1.5) mm. Strobili solitary, 0.5–3.5 cm; sporophylls deltate-ovate to ovate-lanceolate, strongly tapering or not toward apex, abaxial ridges well defined, base glabrous, margins ciliate to slightly dentate, apex slightly keeled, not truncate in profile, long-bristled. 2n = 18.
Habitat: Dry ledges, sea cliffs, limestone, open fire-barrens, sandstone, granite outcrops, exposed rock, rock crevices, sandy or gravelly soil or grassy meadows
Elevation: 0–1900 m
Distribution
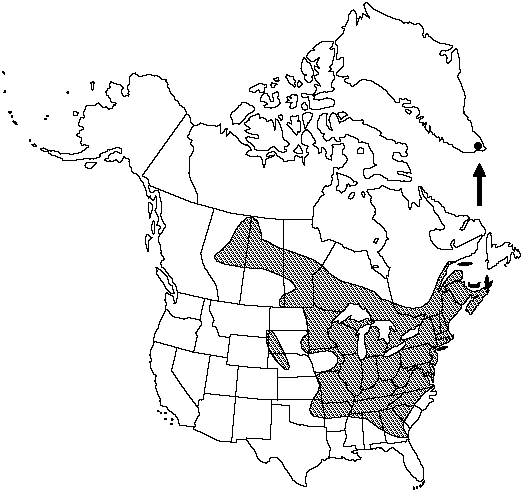
Greenland, Alta., Man., N.B., N.S., Ont., Que., Sask., Ala., Ark., Conn., Del., Ga., Ill., Ind., Iowa., Ky., Kans., Maine, Md., Mass., Mich., Minn., Miss., Mo., Nebr., N.H., N.J., N.Y., N.C., Ohio., Okla., Pa., R.I., S.C., S.Dak., Tenn., Vt., Va., W.Va., Wis., Wyo.
Discussion
Selaginella rupestris has the widest range of any selaginella in the flora. It is variable in many characteristics, e.g., the hairiness of the margins (which sometimes are not hairy), leaf base pubescence, and shape of sporophylls. The variation in sporangial distribution pattern in the strobili and the number of megaspores per megasporangium are important for understanding reproduction and relationships in this species. Very often the strobili are wholly megasporangiate, with only 1–2 megaspores per megasporangium, suggesting asexual reproduction. In other cases, both types of sporangia are present in a strobilus, suggesting sexual reproduction. R. M. Tryon (1971) correlated sporangial and spore distribution patterns in S. rupestris with distributional ranges and concluded that there are four races. Race A has 4 megaspores per megasporangium, has microsporangia, is sexual, and is distributed from southeastern Pennsylvania south to Georgia and Alabama. Race B has 1–2(–4) megaspores per megasporangium, has microsporangia, has an unknown type of reproduction, and has the same range as Race A, but it extends into New York. Race C has 1–2 megaspores per megasporangium, has microsporangia, is probably asexual, and is distributed throughout the species range. Race D has 1–2 megaspores per megasporangium, lacks microsporangia, is therefore asexual, and is found throughout the species range except where Race A occurs. These patterns suggest the presence of more than one species within S. rupestris in the broad sense. More studies, especially cytologic, are needed, as well as fieldwork, in order to understand these relationships and the variability in S. rupestris. Among the species in the flora, S. rupestris seems to be most closely related to S. sibirica (see discussion) and may also be allied to the S. arenicola complex (see discussion).
Selected References
None.