Salix alba
Sp. Pl. 2: 1021. 1753.
Trees, 10–25 m. Stems: branches flexible or ± brittle at base, yellow, gray-brown, or red-brown, glabrous or hairy; branchlets yellowish or gray to red-brown, pilose, densely villous, or long-silky. Leaves: stipules rudimentary or absent on early ones, rudimentary or foliaceous on late ones, apex acute; petiole shallowly grooved adaxially, 3–13 mm, with pairs or clusters of spherical glands or lobes distally, long-silky abaxially; largest medial blade amphistomatous, narrowly oblong, very narrowly elliptic, narrowly elliptic, or lanceolate, 63–115 × 10–20 mm, 4.2–7.3 times as long as wide, base cuneate or convex, margins flat, serrate or serrulate, apex acuminate, caudate, or acute, abaxial surface very densely long-silky to glabrescent, hairs straight, (glaucous or obscured by hairs), adaxial dull, sparsely long-silky; proximal blade margins entire; juvenile blade yellowish green or reddish, very densely long-silky abaxially. Catkins: staminate 27–60 × 6–10 mm, flowering branchlet 2–8 mm; pistillate loosely flowered, slender, 31–51 × 4–8 mm, flowering branchlet 3–14 mm; floral bract 1.6–2.8 mm, apex rounded, entire, abaxially sparsely hairy, hairs straight. Staminate flowers: adaxial nectary oblong to square, 0.3–0.7 mm, nectaries usually distinct (rarely connate); filaments distinct, hairy on proximal 1/2 or basally; anthers (purple, turning yellow), shortly cylindrical to globose, 0.5–0.7 mm. Pistillate flowers: adaxial nectary square, 0.3–0.7 mm, equal to or shorter than stipe; stipe 0.2–0.8 mm; ovary obclavate to pyriform, beak slightly bulged below styles; ovules 8 or 9 per ovary; styles connate, 0.2–0.4 mm; stigmas flat, abaxially non-papillate with rounded tip, or broadly cylindrical, 0.3–0.6 mm. Capsules 3.5–5 mm. 2n = 76.
Phenology: Flowering early May-late Jun.
Habitat: Riverbanks, sandy beaches, fens, old fields, roadsides, gravel pits
Elevation: 70-2000 m
Distribution
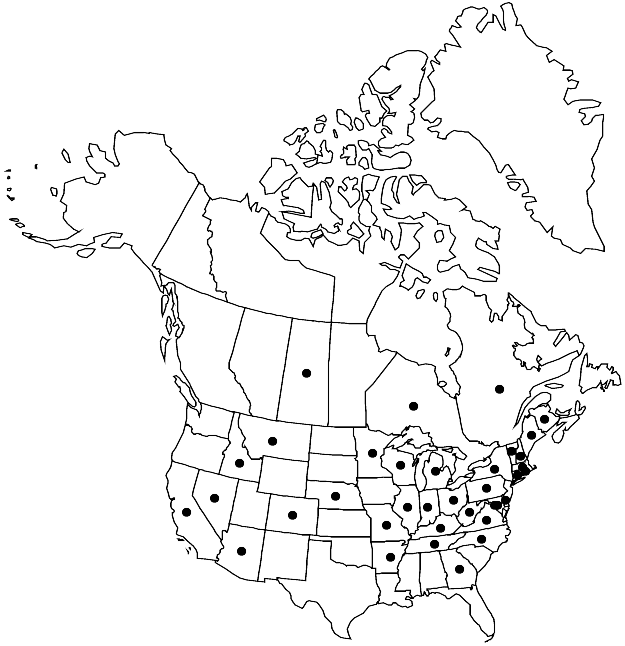
Introduced; N.B., Ont., Que., Sask., Ariz., Ark., Calif., Colo., Conn., Del., D.C., Ga., Idaho, Ill., Ind., Ky., Maine, Md., Mass., Mich., Minn., Mo., Mont., Nebr., Nev., N.H., N.Y., N.C., Ohio, Pa., R.I., Tenn., Vt., Va., W.Va., Wis., Eurasia.
Discussion
Reports that Salix alba is naturalized in Alberta, British Columbia, California, and Washington are undocumented.
The variants of Salix alba, commonly cultivated in the flora area, are often treated as subspecies (K. H. Rechinger 1993) or varieties (R. D. Meikle 1984) but they are all cultivars. The most common ones are: S. alba cv. Sericea (S. alba var. sericea Gaudin) with densely and persistently long-silky leaves and branchlets; S. alba cv. Vitellina (S. alba var. vitellina (Linnaeus) Stokes) with yellow to yellow-brown branchlets and branches; S. alba cv. Caerulea (S. alba var. caerulea (Smith) Smith) with dark brown branchlets and leaves coarsely toothed and sparsely silky abaxially; and S. alba cv. Chermesina (S. alba var. chermesina Hartig) with reddish twigs. Plants referred to in the literature as S. alba var. vitellina cv. Pendula are treated here as S. ×sepulcralis.
Hybrids:
Salix alba forms natural hybrids with S. lucida and S. nigra. Hybrids with S. petiolaris have been reported (M. L. Fernald 1950) but no convincing specimens have been seen.
Salix alba × S. nigra is an infrequent hybrid that is reported to have the catkins of S. alba and the foliage of S. nigra (M. S. Bebb 1895). Buds of this hybrid should be examined to see if they have the partially connate bud-scales characteristic of crosses between species with connate and distinct margins (J. Chmela 1978).
Salix alba × S. euxina. See S. ×fragilis below.
Salix alba × S. lucida. See S. ×jesupii [p. 43].
Salix alba var. vitellina × S. babylonica. See S. ×sepulcralis Simonkai [p. 40].
Salix ×fragilis Linnaeus: The hybrid white willow, S. alba Linnaeus × S. euxina I. Belyaeva, a European introduction, is the most commonly cultivated and naturalized tree-willow in the flora area. It is characterized by: trees, 3–20 m, stems erect or drooping; branches highly brittle at base; petioles with spherical or foliaceous glands distally, pilose or villous adaxially; largest medial leaf blade amphistomatous, very narrowly elliptic or narrowly elliptic, margins uniformly serrate or serrulate, abaxial surface glaucous, both surfaces sparsely long-silky to glabrescent, adaxial surface slightly glossy or dull; juvenile leaves at first densely long-silky soon glabrous; pistillate bract deciduous after flowering; stamens 2; anthers yellow; pistillate adaxial nectary shorter than or equal to stipe; stipe 0.3–0.5 mm; ovary pyriform, glabrous; ovules 6–12 per ovary; styles 0.4–1 mm; capsules 4.5–6 mm; 2n = 57, 76. Flowering is in late May–early June. Individual trees can persist for years by trunk suckering and spread vegetatively by shoot fragmentation along stream margins, shingle and sand beaches, sedge meadows, hardwood forests, and sand pits. It occurs from 0 to 2500 m in Alberta, British Columbia, Manitoba, New Brunswick, Newfoundland, Nova Scotia, Ontario, Prince Edward Island, Quebec, Saskatchewan; Alaska, Arizona, Arkansas, California, Colorado, Connecticut, Delaware, District of Columbia, Georgia, Idaho, Illinois, Indiana, Iowa, Kentucky, Maine, Maryland, Massachusetts, Michigan, Minnesota, Missouri, Montana, Nebraska, Nevada, New Hampshire, New Jersey, New Mexico, New York, Ohio, Oregon, Pennsylvania, Rhode Island, South Dakota, Tennessee, Utah, Vermont, Virginia, Washington, West Virginia, Wisconsin, and Wyoming.
A study of Salix ×fragilis in Colorado (as S. ×rubens) showed that 2172 of 2175 trees were pistillate. Occasionally seed was set, possibly fertilized by S. alba (P. B. Shafroth et al. 1994). There are at least five clones of S. ×fragilis (as S. ×rubens) in cultivation (T. Berg in B. Jonsell and T. Karlsson 2000+, vol. 1); the pistillate are sterile but the staminate produce viable pollen. The hybrid plants are often misidentified as S. “fragilis” or as S. nigra. In the flora area, reproduction of the hybrid seems to be mainly by stem fragmentation.
Prior to the lectotypification of Salix fragilis Linnaeus and the description of S. euxina (I. V. Belyaeva 2009), the name S. “fragilis” was often inadvertently used for both the pure species and for its hybrids with S. alba. Thus all herbarium specimens under the names “fragilis” and “×rubens” need to be revised.
Salix ×fragilis can be separated from S. euxina by having branches and branchlets hairy or glabrescent in age versus glabrous; leaf blades not glaucous abaxially versus glaucous; leaves amphistomatous versus hypostomatous or with stomata only along veins and at apex; and pistillate catkins slender and loosely flowered versus stout and moderately densely flowered.
Several molecular studies have been designed to understand the nature of this hybrid. H. Beissmann et al. (1997), using AFLP markers, were able to recognize three clusters: Salix alba, S. euxina (as S. fragilis), and S. ×fragilis (as S. ×rubens); but a study by K. De Cock et al. (2003), also using AFLP markers, was unable to resolve S. alba and S. ×fragilis (as S. ×rubens). They recommended the use of experimental hybridization to study the genesis of this hybrid. Molecular and genetic studies by L. L. Triest (2001) and coworkers concluded that in modern open agricultural situations in Belgium, hybridization was of low occurrence, and that morphologically intermediate plants were not necessarily genetically intermediate. These studies saw different facets of the question. Clearly there are three entities, S. alba, S. euxina, and their hybrid but, because S. euxina may be rare outside of cultivation, natural hybridization may not occur and the question of whether S. ×fragilis can be backcrossed with S. alba remains to be studied. The specimens used in these molecular studies require reidentification.
Salix ×jesupii Fernald: S. alba × S. lucida; S. ×ehrhartiana of authors, not G. Meyer. The origin of this hybrid, between a European and a native species, is unknown. It is characterized by: shrubs or trees, 7–10 m; branchlets red-brown; petioles convex to shallowly grooved adaxially, 3–11 mm, with pairs or clusters of spherical or stalked glands distally; largest medial leaf blade amphistomatous or hemiamphistomatous, abaxial surface glaucous, sparsely long-silky to glabrescent, hairs white and ferruginous; floral bract apex acute to rounded, toothed or entire; pistillate flowering branchlet 8–16 mm, bract deciduous after flowering; stamens 3–5; stipes 0.3–0.6 mm; ovary pyriform, glabrous; ovules 12–14 per ovary; styles 0.3–0.8 mm; capsules 4–5 mm. Flowering is late April–mid May. It occurs in the flora area on the edges of streams and lakes, in wet deciduous woods, sand dunes, and wet railroad rights-of-way, at 10–300 m, in Ontario, Quebec, Prince Edward Island, Saskatchewan; Illinois, Kentucky, Massachusetts, New Hampshire, New York, Ohio, Pennsylvania, Vermont, Virginia, Washington, West Virginia, and Wisconsin. It is possibly introduced in Washington.
Salix ×jesupii was named by M. S. Bebb (1895) as a formula hybrid, S. alba × S. lucida, and Fernald based his binomial on Bebb’s exsiccatae. In North America, it was mistaken for the European S. ×ehrhartiana G. Meyer (S. alba Linnaeus × S. pentandra Linnaeus) (G. W. Argus 1986). Study of nectary morphology confirmed that the North American plants were not the same as the European hybrid. In herbaria, plants of S. ×jesupii often are misidentified as S. alba, S. euxina, S. ×fragilis, or S. lucida.
Selected References
None.