Cystopteris tennesseensis
J. Tennessee Acad. Sci. 25(2): 107. 1950.
Stems creeping, not cordlike, internodes short, heavily beset with old petiole bases, hairs absent; scales usually tan to light brown, lanceolate, radial walls tan to brown, thin, luminae tan. Leaves monomorphic, crowded near stem apex, to 45 cm, nearly all bearing sori. Petiole variable in color but mostly dark brown at base, gradually becoming straw-colored distally, shorter than blade, sparsely scaly at base. Blade deltate to narrowly deltate, 2-pinnate-pinnatifid, usually widest at or near base, apex short-attenuate; rachis and costae with occasional unicellular, gland-tipped hairs, with or without bulblets (usually misshapen); axils of pinnae with infrequent multicellular, gland-tipped hairs. Pinnae usually perpendicular to rachis, not curving toward blade apex, margins serrate; proximal pinnae pinnatifid to pinnate-pinnatifid, ± equilateral, basiscopic pinnules not enlarged, basal basiscopic pinnules sessile to short-stalked, base truncate to obtuse; distal pinnae ovate to oblong. Veins directed into teeth and notches. Indusia cup-shaped, apex truncate, with scattered, unicellular, gland-tipped hairs. Spores spiny, usually 38–42 µm. 2n = 168.
Phenology: Sporulating summer–fall.
Habitat: Cracks and ledges on cliffs, rarely terrestrial, often on calcareous substrates or associated with man-made habitats such as rock walls or bridge abutments
Elevation: 100–500 m
Distribution
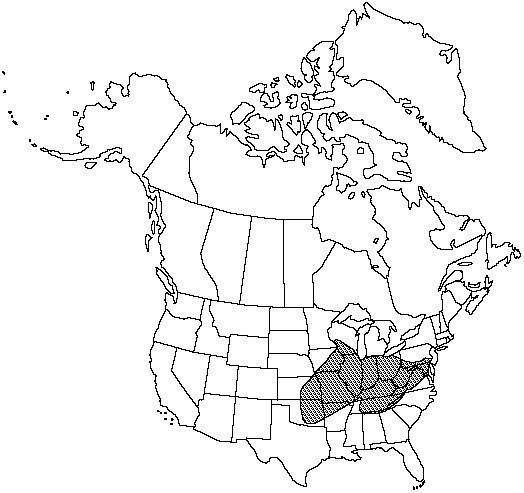
Ala., Ark., Ga., Ill., Ind., Iowa, Kans., Ky., Md., Mo., N.C., Ohio., Okla., Pa., Tenn., Va., W.Va., Wis.
Discussion
Cystopteris tennesseensis, an allotetraploid species, has C. bulbifera and C. protrusa as diploid progenitors. The relative distinctiveness of these diploids suggests that identification of C. tennesseensis individuals should be straightforward. As with other members of Cystopteris, however, a series of features makes reliable recognition of this tetraploid challenging. For some characteristics (occasional unicellular, gland-tipped hairs and bulblets; short-attenuate, narrowly deltate blades), it is intermediate between its parents; for others (very short internodes and crowded leaves; occurrence on rock), it tends toward C. bulbifera. This unequal intermediacy, the multiple origins from genetically different individuals (C. H. Haufler et al. 1990), and the occurrence of sterile backcross triploids with its diploid progenitors in zones of sympatry has blurred the already subtle features distinguishing this allopolyploid. For example, some individuals of C. bulbifera may have very few glandular hairs, and some C. tennesseensis appear to lack glandular hairs entirely (R. F. Blasdell 1963). Further, sterile tetraploid hybrids (called C. × wagneri R. C. Moran) between C. tennesseensis and C. tenuis have been reported (R. C. Moran 1983) and verified through isozyme analyses (C. H. Haufler, unpubl. data). Finally, as discussed above, the recently recognized C. utahensis (C. H. Haufler and M. D. Windham 1991) is extremely similar morphologically to C. tennesseensis.
Selected References
None.