Amelanchier arborea
Rhodora 43: 563. 1941.
Shrubs or trees, 2–20 m. Stems 1–20, solitary or in colonies. Leaves less than half-unfolded; petiole (5.5–)12–20.6(–29.2) mm; blade ovate to obovate, (30–)47–68(–93) × (16–)26–40(–56) cm, base cordate to rounded, each margin with (0–)11–21(–30) teeth on proximal 1/2 and (3–)5–9(–13) teeth in distalmost cm, largest teeth less than 1 mm, apex acute to acuminate, abaxial surface densely (moderately) hairy by flowering, glabrous or sparsely to moderately hairy later, adaxial glabrous or sparsely (moderately) hairy later. Inflorescences (3–)6–12(–15)-flowered, (19–)30–55(–79) mm. Pedicels: 0 or 1 subtended by a leaf, proximalmost (5–)10–20(–32) mm. Flowers: sepals soon reflexed after flowering, (1.6–)2.3–3.7(–5.3) mm; petals linear to oblong, (8–)10–15(–19) × (2–)3.1–5(–6.8) mm; stamens (16–)20(–21); styles (3–)5(or 6), (2.1–)3.2–4.2(–5.5) mm; ovary apex glabrous or sparsely (densely) hairy. Pomes maroon-purple, 6–10 mm diam. 2n = 2x, 4x.
Phenology: Flowering Feb–May; fruiting May–Jul.
Habitat: Dry to moist woods, mesic mixed hardwoods and pine-hardwoods, fields, thickets, roadsides, circumneutral soil, especially northward
Elevation: 0–1500 m
Distribution
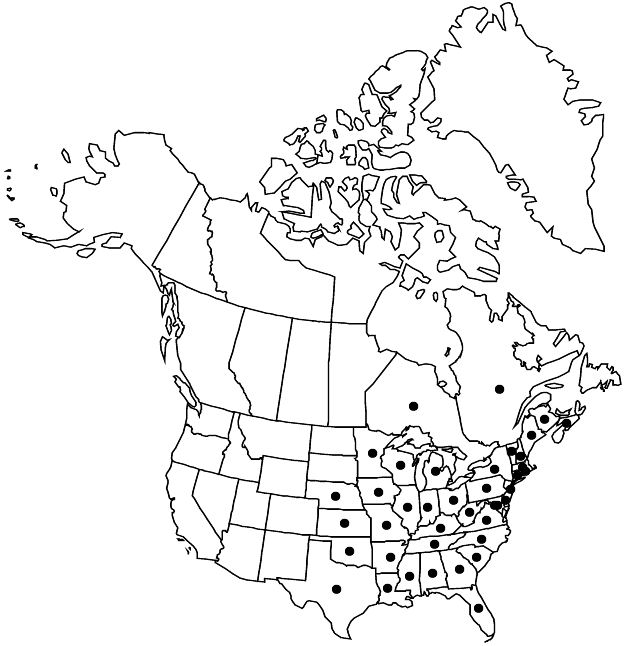
N.B., N.S., Ont., Que., Ala., Ark., Conn., Del., D.C., Fla., Ga., Ill., Ind., Iowa, Kans., Ky., La., Maine, Md., Mass., Mich., Minn., Miss., Mo., Nebr., N.H., N.J., N.Y., N.C., Ohio, Okla., Pa., R.I., S.C., Tenn., Tex., Vt., Va., W.Va., Wis.
Discussion
Amelanchier arborea is distinctive for its arborescent habit, generally precocious flowering, leaf margins each with (3–)5–9(–13) teeth on distal cm, densely hairy abaxial surface of leaf blades, and strongly reflexed sepals at flowering. Leaves typically have subcordate to strongly cordate bases, acuminate apices, and margins sometimes finely double serrate.
The name Amelanchier canadensis was applied by many botanists to A. arborea and A. laevis until M. L. Fernald (1941) clarified the issue; this usage appears in some older manuals. McKay transferred A. laevis to subspecific status under A. arborea, and the two species are generally considered to be closely related. Allozyme data show that, with one exception, populations of each are more closely related to a nearby population of the other than they are to conspecific populations (R. D. Overath and J. L. Hamrick 1998). At flowering, A. arborea and A. laevis can be distinguished by leaf color, which is typically reddish green or reddish brown in A. laevis and green to greenish white in A. arborea. In addition, the leaves of A. laevis are at least half-expanded and unfolding as well as glabrous (or sparsely hairy) by flowering, and its sepals are spreading to reflexed.
Amelanchier arborea exhibits considerable variation; infraspecific taxa have been named. Variety alabamensis differs in its densely hairy ovary apices. Variation in ovary summit indument in A. arborea does not appear to be associated with any other taxonomically informative variation; individuals with varying amounts of ovary indument occur sporadically in populations; this does not appear ecologically significant. The authors conclude that this variety does not warrant taxonomic recognition; they have observed groups of populations that differ in the number of flowers per inflorescence, color of the expanding leaves, size of the teeth, and other features.
Amelanchier arborea usually flowers a week or more before sympatric congeners. The species has been reported (M. L. Fernald 1950) to hybridize with A. bartramiana, A. canadensis, A. humilis, and A. laevis. The authors have collected putative hybrids with A. amabilis in New York and have observed putative hybrids between A. arborea and other eastern North American species of Amelanchier. Hybrid swarms in New Jersey between this species and A. laevis but not A. canadensis have been reported (J. E. Cruise 1964).
Selected References
None.