Difference between revisions of "Cheilanthes bonariensis"
Bull. Inst. Jamaica, Sci. Ser. 5(1): 15. 1953.
FNA>Volume Importer |
imported>Volume Importer |
||
| (7 intermediate revisions by 2 users not shown) | |||
| Line 8: | Line 8: | ||
}} | }} | ||
|common_names=Bonaire lip fern | |common_names=Bonaire lip fern | ||
| − | |basionyms={{Treatment/ID/ | + | |basionyms={{Treatment/ID/Basionym |
|name=Acrostichum bonariense | |name=Acrostichum bonariense | ||
|authority=Willdenow | |authority=Willdenow | ||
| + | |rank=species | ||
| + | |publication_title=Sp. Pl. | ||
| + | |publication_place=5: 114. 1810 | ||
}} | }} | ||
|synonyms={{Treatment/ID/Synonym | |synonyms={{Treatment/ID/Synonym | ||
|name=Notholaena aurea | |name=Notholaena aurea | ||
|authority=(Poiret) Desvaux | |authority=(Poiret) Desvaux | ||
| + | |rank=species | ||
}} | }} | ||
|hierarchy=Pteridaceae;Cheilanthes;Cheilanthes bonariensis | |hierarchy=Pteridaceae;Cheilanthes;Cheilanthes bonariensis | ||
| Line 23: | Line 27: | ||
}}<!-- | }}<!-- | ||
| − | --><span class="statement" id="st- | + | --><span class="statement" id="st-undefined" data-properties=""><b>Stems </b>short-creeping to compact, usually 4–8 mm diam.; scales bicolored, with broad, well-defined, dark, central stripe and narrow, light brown margins, narrowly lanceolate, slightly contorted, strongly appressed, persistent. <b>Leaves</b> clustered, 10–60 cm; vernation noncircinate. <b>Petiole</b> dark brown, rounded adaxially. <b>Blade</b> linear, pinnate-pinnatifid throughout, 1–4 cm wide; rachis rounded adaxially, lacking scales, with dense monomorphic pubescence. <b>Pinnae</b> articulate at swollen, hirsute nodes, basal pair slightly smaller than adjacent pair, ± equilateral, appearing hirsute adaxially. <b>Costae</b> absent. <b>Ultimate</b> segments elongate-deltate to ovate, not especially beadlike, the largest 1–7 mm, abaxially densely tomentose, adaxially hirsute. <b>False</b> indusia marginal, weakly differentiated, 0.05–0.25 mm wide. <b>Sori</b> ± continuous around segment margins. <b>Sporangia</b> containing 32 spores. n = 2n = 90, apogamous.</span><!-- |
-->{{Treatment/Body | -->{{Treatment/Body | ||
| + | |phenology=Sporulating summer–fall. | ||
|habitat=Rocky slopes and ledges, found on a variety of substrates though rarely observed on limestone | |habitat=Rocky slopes and ledges, found on a variety of substrates though rarely observed on limestone | ||
|elevation=1200–2400 m | |elevation=1200–2400 m | ||
|distribution=Ariz.;N.Mex.;Tex.;Mexico;West Indies;Central America;South America. | |distribution=Ariz.;N.Mex.;Tex.;Mexico;West Indies;Central America;South America. | ||
| − | |discussion=<p>Cheilanthes bonariensis has been assigned to Notholaena in past treatments. It is distantly related (at best) to the species here included in Notholaena, however, and we concur with R. M. Tryon and A. F. Tryon (1982) that it should be transferred to Cheilanthes. Chromosomal studies (G. J. Gastony and M. D. Windham 1989) suggest that C. bonariensis is an apogamous triploid that arose through autopolyploidy. Further investigation is necessary to determine whether 64-spored, sexually reproducing populations of C. bonariensis are still extant.</p> | + | |discussion=<p><i>Cheilanthes bonariensis</i> has been assigned to <i>Notholaena</i> in past treatments. It is distantly related (at best) to the species here included in <i>Notholaena</i>, however, and we concur with R. M. Tryon and A. F. Tryon (1982) that it should be transferred to <i>Cheilanthes</i>. Chromosomal studies (G. J. Gastony and M. D. Windham 1989) suggest that <i>C. bonariensis</i> is an apogamous triploid that arose through autopolyploidy. Further investigation is necessary to determine whether 64-spored, sexually reproducing populations of <i>C. bonariensis</i> are still extant.</p> |
|tables= | |tables= | ||
|references= | |references= | ||
| Line 38: | Line 43: | ||
-->{{#Taxon: | -->{{#Taxon: | ||
name=Cheilanthes bonariensis | name=Cheilanthes bonariensis | ||
| − | |||
|authority=(Willdenow) Proctor | |authority=(Willdenow) Proctor | ||
|rank=species | |rank=species | ||
| Line 45: | Line 49: | ||
|basionyms=Acrostichum bonariense | |basionyms=Acrostichum bonariense | ||
|family=Pteridaceae | |family=Pteridaceae | ||
| + | |phenology=Sporulating summer–fall. | ||
|habitat=Rocky slopes and ledges, found on a variety of substrates though rarely observed on limestone | |habitat=Rocky slopes and ledges, found on a variety of substrates though rarely observed on limestone | ||
|elevation=1200–2400 m | |elevation=1200–2400 m | ||
| Line 52: | Line 57: | ||
|publication year=1953 | |publication year=1953 | ||
|special status= | |special status= | ||
| − | |source xml=https:// | + | |source xml=https://bitbucket.org/aafc-mbb/fna-data-curation/src/2e0870ddd59836b60bcf96646a41e87ea5a5943a/coarse_grained_fna_xml/V2/V2_360.xml |
|genus=Cheilanthes | |genus=Cheilanthes | ||
|species=Cheilanthes bonariensis | |species=Cheilanthes bonariensis | ||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
}}<!-- | }}<!-- | ||
-->[[Category:Treatment]][[Category:Cheilanthes]] | -->[[Category:Treatment]][[Category:Cheilanthes]] | ||
Latest revision as of 21:22, 5 November 2020
Stems short-creeping to compact, usually 4–8 mm diam.; scales bicolored, with broad, well-defined, dark, central stripe and narrow, light brown margins, narrowly lanceolate, slightly contorted, strongly appressed, persistent. Leaves clustered, 10–60 cm; vernation noncircinate. Petiole dark brown, rounded adaxially. Blade linear, pinnate-pinnatifid throughout, 1–4 cm wide; rachis rounded adaxially, lacking scales, with dense monomorphic pubescence. Pinnae articulate at swollen, hirsute nodes, basal pair slightly smaller than adjacent pair, ± equilateral, appearing hirsute adaxially. Costae absent. Ultimate segments elongate-deltate to ovate, not especially beadlike, the largest 1–7 mm, abaxially densely tomentose, adaxially hirsute. False indusia marginal, weakly differentiated, 0.05–0.25 mm wide. Sori ± continuous around segment margins. Sporangia containing 32 spores. n = 2n = 90, apogamous.
Phenology: Sporulating summer–fall.
Habitat: Rocky slopes and ledges, found on a variety of substrates though rarely observed on limestone
Elevation: 1200–2400 m
Distribution
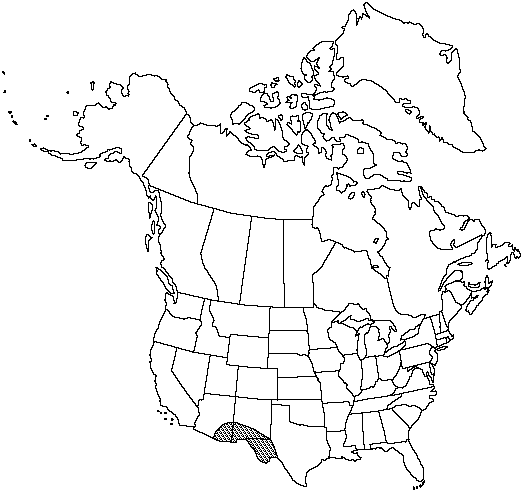
Ariz., N.Mex., Tex., Mexico, West Indies, Central America, South America.
Discussion
Cheilanthes bonariensis has been assigned to Notholaena in past treatments. It is distantly related (at best) to the species here included in Notholaena, however, and we concur with R. M. Tryon and A. F. Tryon (1982) that it should be transferred to Cheilanthes. Chromosomal studies (G. J. Gastony and M. D. Windham 1989) suggest that C. bonariensis is an apogamous triploid that arose through autopolyploidy. Further investigation is necessary to determine whether 64-spored, sexually reproducing populations of C. bonariensis are still extant.
Selected References
None.