Difference between revisions of "Carex gynocrates"
Naturhist. Tidsskr. 3: 434. 1841.
FNA>Volume Importer |
FNA>Volume Importer |
||
| Line 12: | Line 12: | ||
|name=Carex dioica subsp. gynocrates | |name=Carex dioica subsp. gynocrates | ||
|authority=(Wormskjöld ex Drejer) Hultén | |authority=(Wormskjöld ex Drejer) Hultén | ||
| + | |rank=subspecies | ||
}} {{Treatment/ID/Synonym | }} {{Treatment/ID/Synonym | ||
|name=Carex dioica var. gynocrates | |name=Carex dioica var. gynocrates | ||
|authority=(Wormskjöld ex Drejer) Ostenfeld | |authority=(Wormskjöld ex Drejer) Ostenfeld | ||
| + | |rank=variety | ||
}} | }} | ||
|hierarchy=Cyperaceae;Carex;Carex sect. Physoglochin;Carex gynocrates | |hierarchy=Cyperaceae;Carex;Carex sect. Physoglochin;Carex gynocrates | ||
| Line 31: | Line 33: | ||
|distribution=Greenland;St. Pierre and Miquelon;Alta.;B.C.;Man.;N.B.;Nfld. and Labr.;N.W.T.;N.S.;Nunavut;Ont.;Que.;Sask.;Yukon;Alaska;Colo.;Idaho;Maine;Mich.;Minn.;Mont.;Nev.;N.Y.;N.Dak.;Oreg.;Pa.;Utah;Wis.;Wyo.;Asia (e Siberia). | |distribution=Greenland;St. Pierre and Miquelon;Alta.;B.C.;Man.;N.B.;Nfld. and Labr.;N.W.T.;N.S.;Nunavut;Ont.;Que.;Sask.;Yukon;Alaska;Colo.;Idaho;Maine;Mich.;Minn.;Mont.;Nev.;N.Y.;N.Dak.;Oreg.;Pa.;Utah;Wis.;Wyo.;Asia (e Siberia). | ||
|discussion=<p>Chiefly boreal, <i>Carex gynocrates</i> is infrequent to rare southward, especially in the western cordillera. The only Pennsylvania collection was made by Goldie at Pittsburgh between 1819 and 1822.</p><!-- | |discussion=<p>Chiefly boreal, <i>Carex gynocrates</i> is infrequent to rare southward, especially in the western cordillera. The only Pennsylvania collection was made by Goldie at Pittsburgh between 1819 and 1822.</p><!-- | ||
| − | --><p>Opinions differ about the taxonomic rank of the North American and east Asian (Siberian) populations, which have chromosome counts of 2n = 48, 50, and 70, and perhaps are best segregated as <i>Carex</i> dioica subsp. gynocrates (E. Hultén 1962). The latter is only weakly distinguished from the Eurasian < | + | --><p>Opinions differ about the taxonomic rank of the North American and east Asian (Siberian) populations, which have chromosome counts of 2n = 48, 50, and 70, and perhaps are best segregated as <i>Carex</i> dioica subsp. gynocrates (E. Hultén 1962). The latter is only weakly distinguished from the Eurasian <i></i>subsp.<i> dioica</i>, which has chromosome counts of 2n = 52 and 60, and tends to be less strongly dioecious, and has paler pistillate scales and ventrally more convex perigynia.</p><!-- |
--><p>The hybrid between <i>Carex gynocrates</i> and <i>C. maritima</i>, C. ×langeana Fernald (pro sp.) [C. dutillyi O’Neill & Duman], strongly resembles <i>C. maritima</i>; the leaves are slightly scabrous-roughened toward the apex; the heads are smaller, ellipsoid to ovoid-oblong and only 3–5 mm thick; the perigynia are appressed-ascending and ± flat; and the achenes are not well-developed. <i>Carex</i> ×langeana is reported as forming close turf on dry, peaty limestone barrens in Newfoundland (M. L. Fernald 1933, 1950) and as occasional upon humid rocks and coastal fens along Hudson Bay (J. Deshaye and J. Cayouette 1988). The hybrid should be expected where the parents coexist and has been reported from most of those areas (J. Cayouette and P. M. Catling 1992), although the report of the hybrid from Alaska by H. J. Scoggan (1978–1979) is based on a specimen of <i>C. maritima</i>.</p> | --><p>The hybrid between <i>Carex gynocrates</i> and <i>C. maritima</i>, C. ×langeana Fernald (pro sp.) [C. dutillyi O’Neill & Duman], strongly resembles <i>C. maritima</i>; the leaves are slightly scabrous-roughened toward the apex; the heads are smaller, ellipsoid to ovoid-oblong and only 3–5 mm thick; the perigynia are appressed-ascending and ± flat; and the achenes are not well-developed. <i>Carex</i> ×langeana is reported as forming close turf on dry, peaty limestone barrens in Newfoundland (M. L. Fernald 1933, 1950) and as occasional upon humid rocks and coastal fens along Hudson Bay (J. Deshaye and J. Cayouette 1988). The hybrid should be expected where the parents coexist and has been reported from most of those areas (J. Cayouette and P. M. Catling 1992), although the report of the hybrid from Alaska by H. J. Scoggan (1978–1979) is based on a specimen of <i>C. maritima</i>.</p> | ||
|tables= | |tables= | ||
| Line 41: | Line 43: | ||
-->{{#Taxon: | -->{{#Taxon: | ||
name=Carex gynocrates | name=Carex gynocrates | ||
| − | |||
|authority=Wormskjöld ex Drejer | |authority=Wormskjöld ex Drejer | ||
|rank=species | |rank=species | ||
| Line 56: | Line 57: | ||
|publication year=1841 | |publication year=1841 | ||
|special status= | |special status= | ||
| − | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/ | + | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/f50eec43f223ca0e34566be0b046453a0960e173/coarse_grained_fna_xml/V23/V23_523.xml |
|genus=Carex | |genus=Carex | ||
|section=Carex sect. Physoglochin | |section=Carex sect. Physoglochin | ||
Revision as of 20:09, 16 December 2019

Rhizomes horizontal, threadlike, 0.3–0.8 mm in diam. Culms arising singly or 2–3 together, ± terete, scarcely furrowed, 2–30 cm; lateral sterile shoots often with prostrate or curved base. Leaf blades filiform, 2–15 cm × 0.3–0.7 mm. Inflorescence androgynous (14%), wholly staminate (12%), or wholly pistillate (74%); staminate spike 8–16 mm; pistillate spike ± densely 4–15(–18)-flowered, transversely broadly oblong to ovoid-oblong or oblong, 5–14 × 4–8 mm. Pistillate scales uniformly light to dark brown or with pale or green midvein, margins scarious, narrow to broad. Perigynia becoming divergent or slightly deflexed, yellow, olive, or at maturity chestnut-brown, obscurely to evidently 17–20-striate (marginal ribs seldom prominent), oblong-ovoid, biconvex, 2.9–3.4 × 1.2–1.7 mm; beak 0.5 mm, margins glabrous or sparsely scabrid. Achenes 1.5–1.7 × 1–1.2 mm. 2n = 46, 48.
Phenology: Fruiting summer (Jun–Aug, Sep in subarctic Quebec and Greenland).
Habitat: Wet peaty ground, usually in openings in coniferous swamps and conifer-hardwood stands, less often in poor fens, boggy swales (flarks) and alder thickets, also subalpine meadows, tundra, outwash gravel and seepage areas, usually on calcareous substrates
Elevation: 0–3100 m
Distribution
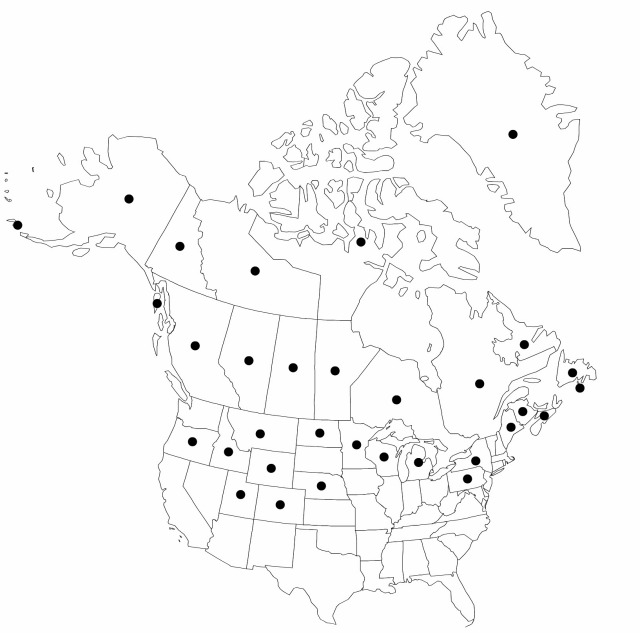
Greenland, St. Pierre and Miquelon, Alta., B.C., Man., N.B., Nfld. and Labr., N.W.T., N.S., Nunavut, Ont., Que., Sask., Yukon, Alaska, Colo., Idaho, Maine, Mich., Minn., Mont., Nev., N.Y., N.Dak., Oreg., Pa., Utah, Wis., Wyo., Asia (e Siberia).
Discussion
Chiefly boreal, Carex gynocrates is infrequent to rare southward, especially in the western cordillera. The only Pennsylvania collection was made by Goldie at Pittsburgh between 1819 and 1822.
Opinions differ about the taxonomic rank of the North American and east Asian (Siberian) populations, which have chromosome counts of 2n = 48, 50, and 70, and perhaps are best segregated as Carex dioica subsp. gynocrates (E. Hultén 1962). The latter is only weakly distinguished from the Eurasian subsp. dioica, which has chromosome counts of 2n = 52 and 60, and tends to be less strongly dioecious, and has paler pistillate scales and ventrally more convex perigynia.
The hybrid between Carex gynocrates and C. maritima, C. ×langeana Fernald (pro sp.) [C. dutillyi O’Neill & Duman], strongly resembles C. maritima; the leaves are slightly scabrous-roughened toward the apex; the heads are smaller, ellipsoid to ovoid-oblong and only 3–5 mm thick; the perigynia are appressed-ascending and ± flat; and the achenes are not well-developed. Carex ×langeana is reported as forming close turf on dry, peaty limestone barrens in Newfoundland (M. L. Fernald 1933, 1950) and as occasional upon humid rocks and coastal fens along Hudson Bay (J. Deshaye and J. Cayouette 1988). The hybrid should be expected where the parents coexist and has been reported from most of those areas (J. Cayouette and P. M. Catling 1992), although the report of the hybrid from Alaska by H. J. Scoggan (1978–1979) is based on a specimen of C. maritima.
Selected References
None.