Difference between revisions of "Rosa woodsii subsp. manca"
Novon 17: 351. 2007.
FNA>Volume Importer |
FNA>Volume Importer |
||
| Line 34: | Line 34: | ||
|elevation=2100–3300 m | |elevation=2100–3300 m | ||
|distribution=Ariz.;Colo.;N.Mex.;Utah;Wyo. | |distribution=Ariz.;Colo.;N.Mex.;Utah;Wyo. | ||
| − | |discussion=<p>Subspecies manca [2x (DNA)] is the high-elevation representative of Rosa woodsii in the southern Rocky Mountains and outlying peaks and ridges of the Colorado Plateau, particularly characteristic of Gambel oak woodlands. Plants are relatively short and tend to have curved prickles, obovate leaflets, and stipitate-glandular sepals. As so circumscribed, this is not the R. manca of E. W. Erlanson (1934), which was regarded as a hexaploid (2n = 42) ecotype of R. nutkana. Neither is it the R. manca of A. Cronquist and N. H. Holmgren (1997), which represented a mixture of subsp. manca and R. nutkana, probably subsp. melina.</p> | + | |discussion=<p>Subspecies manca [2x (DNA)] is the high-elevation representative of <i>Rosa woodsii</i> in the southern Rocky Mountains and outlying peaks and ridges of the Colorado Plateau, particularly characteristic of Gambel oak woodlands. Plants are relatively short and tend to have curved prickles, obovate leaflets, and stipitate-glandular sepals. As so circumscribed, this is not the R. manca of E. W. Erlanson (1934), which was regarded as a hexaploid (2n = 42) ecotype of <i>R. nutkana</i>. Neither is it the R. manca of A. Cronquist and N. H. Holmgren (1997), which represented a mixture of <i></i>subsp.<i> manca</i> and <i>R. nutkana</i>, probably <i></i>subsp.<i> melina</i>.</p> |
|tables= | |tables= | ||
|references= | |references= | ||
| Line 58: | Line 58: | ||
|publication year=2007 | |publication year=2007 | ||
|special status=Endemic;Selected by author to be illustrated | |special status=Endemic;Selected by author to be illustrated | ||
| − | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/ | + | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/8f726806613d60c220dc4493de13607dd3150896/coarse_grained_fna_xml/V9/V9_156.xml |
|subfamily=Rosaceae subfam. Rosoideae | |subfamily=Rosaceae subfam. Rosoideae | ||
|tribe=Rosaceae tribe Roseae | |tribe=Rosaceae tribe Roseae | ||
Revision as of 18:15, 18 September 2019

Shrubs, 2–10(+) dm. Stems usually densely branched; prickles usually present on distal stems and branches, infrastipular curved or hooked, rarely erect, terete, 3–6 mm, internodal usually dense, rarely sparse or absent. Terminal leaflets obovate, sometimes ovate or elliptic, (6–)13–17(–35) mm. Inflorescences 1–3(+)-flowered. Sepals: abaxial surfaces usually stipitate-glandular.
Phenology: Flowering Jun–Jul.
Habitat: Rocky meadows, high mountain forests to aspen and scrub oak woodlands, along streams, moist ditch banks
Elevation: 2100–3300 m
Distribution
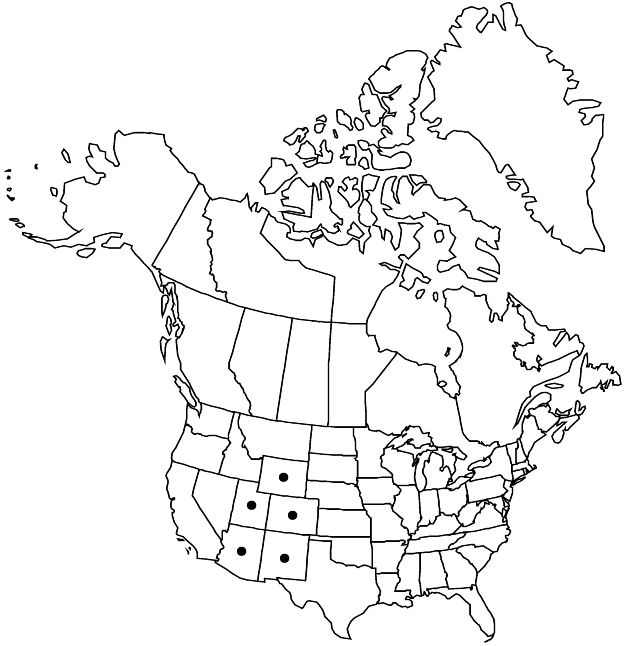
Ariz., Colo., N.Mex., Utah, Wyo.
Discussion
Subspecies manca [2x (DNA)] is the high-elevation representative of Rosa woodsii in the southern Rocky Mountains and outlying peaks and ridges of the Colorado Plateau, particularly characteristic of Gambel oak woodlands. Plants are relatively short and tend to have curved prickles, obovate leaflets, and stipitate-glandular sepals. As so circumscribed, this is not the R. manca of E. W. Erlanson (1934), which was regarded as a hexaploid (2n = 42) ecotype of R. nutkana. Neither is it the R. manca of A. Cronquist and N. H. Holmgren (1997), which represented a mixture of subsp. manca and R. nutkana, probably subsp. melina.
Selected References
None.