Difference between revisions of "Amelanchier interior"
Amer. Midl. Naturalist 22: 185, plate 13. 1939.
FNA>Volume Importer |
imported>Volume Importer |
||
| Line 55: | Line 55: | ||
|publication year=1939 | |publication year=1939 | ||
|special status=Endemic | |special status=Endemic | ||
| − | |source xml=https:// | + | |source xml=https://bibilujan@bitbucket.org/aafc-mbb/fna-data-curation.git/src/bb6b7e3a7de7d3b7888a1ad48c7fd8f5c722d8d6/coarse_grained_fna_xml/V9/V9_1126.xml |
|subfamily=Rosaceae subfam. Amygdaloideae | |subfamily=Rosaceae subfam. Amygdaloideae | ||
|tribe=Rosaceae tribe Gillenieae | |tribe=Rosaceae tribe Gillenieae | ||
Revision as of 00:25, 28 May 2020
Shrubs or trees, 1–10 m. Stems 1–10, forming colonies or solitary. Leaves not fully unfolded; petiole 10–30 mm; blade broadly ovate to elliptic, 30–70 × 20–50 mm, base rounded to subcordate, each margin with 3–15 teeth on proximal 1/2 and (2–)4 or 5(–7) teeth in distalmost cm, largest teeth less than 1 mm, apex short acuminate to apiculate, abaxial surface glabrous or sparsely hairy by flowering, surfaces glabrous later. Inflorescences 4–12-flowered, 30–75 mm. Pedicels: 1 or 2 subtended by a leaf, proximalmost 10–45 mm. Flowers: sepals recurved after flowering, 2–5 mm; petals obovate, 6–15 × 4–5 mm; stamens 20; styles 5, 3–5 mm; ovary apex densely (moderately) hairy (or glabrous). Pomes purple-black, 6–8 mm diam. 2n = 4x.
Phenology: Flowering May–Jun; fruiting Jul–Aug.
Habitat: Dry woods, bluffs rocky areas and slopes, stream banks, fields, thickets, and sandy areas, sometimes wetlands
Elevation: 0–300 m
Distribution
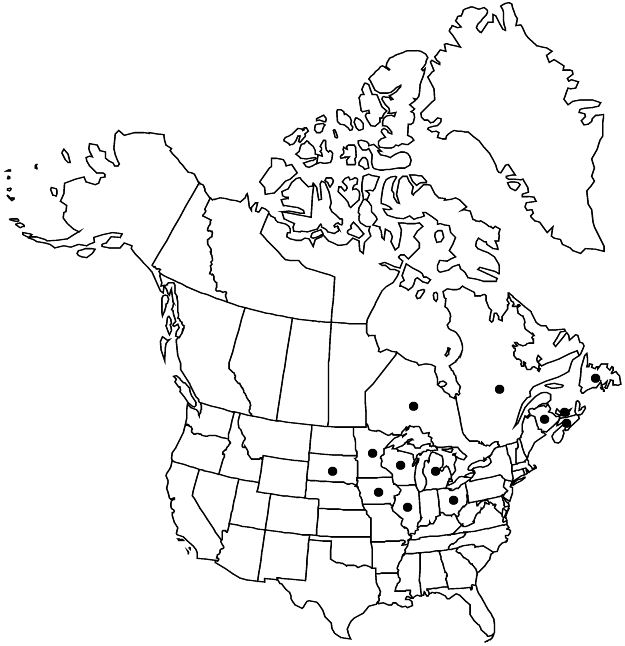
N.B., Nfld. and Labr. (Nfld.), N.S., Ont., P.E.I., Que., Ill., Iowa, Mich., Minn., Ohio, S.Dak., Wis.
Discussion
Amelanchier interior is distinguished by its capacity to grow into a tree to ten meters and by having sparsely hairy young leaves that are often reddish and densely hairy ovary apices. E. L. Nielsen (1939) differentiated A. interior and A. wiegandii on the basis of leaves being carinate in A. wiegandii as opposed to flat in A. interior, and leaf sinuses rounded versus acute. G. N. Jones (1946) considered those differences to be slight, and he included A. wiegandii in A. interior. The authors follow Jones, but retain the common name Wiegand’s shadbush to commemorate Wiegand’s early insights about the taxonomy of Amelanchier. M. L. Fernald (1950) considered A. wiegandii as suggesting a small-leaved A. laevis but with shorter buds, fewer teeth, fewer veins, and summit of ovary heavily tomentose. P. Landry (1975) thought A. wiegandii to be the hybrid of A. arborea and A. sanguinea, and E. G. Voss (1972–1996, vol. 2) reasoned that A. interior is a hybrid swarm involving A. laevis (or sometimes A. arborea) and plants of the A. spicata and/or A. sanguinea complex. A hybrid origin of A. interior from A. laevis and A. sanguinea is reasonable given that stem height, the number of leaf teeth, and petal length are more or less intermediate between those two species. DNA sequences from ITS region indicate that A. interior (A. wiegandii) is a possible later-generation hybrid involving a member of the western North American ITS clade (which also includes A. humilis and A. sanguinea of eastern North America) and some eastern North American taxon (C. S. Campbell et al. 1997). Multiple hybrid origins, possibly from different species, may explain the variability of A. interior. Amelanchier interior has unusually large ranges of lengths for proximal pedicels, sepals, and petals. The leaves of A. wiegandii were described by Nielsen as bronze at flowering; Jones described the leaves of A. interior as green when young. The authors assume that A. interior as they interpret it is polymorphic for the color of young leaves.
Selected References
None.