Difference between revisions of "Gymnocarpium dryopteris"
Phytologist 4: app. 24. 1851.
FNA>Volume Importer |
GeoffLevin (talk | contribs) m (Fixed Nfld. and Labr. distribution to match printed version.) |
||
| (8 intermediate revisions by 2 users not shown) | |||
| Line 8: | Line 8: | ||
}} | }} | ||
|common_names=Common oak fern;gymnocarpe fougère-du-chêne | |common_names=Common oak fern;gymnocarpe fougère-du-chêne | ||
| − | |basionyms={{Treatment/ID/ | + | |basionyms={{Treatment/ID/Basionym |
|name=Polypodium dryopteris | |name=Polypodium dryopteris | ||
|authority=Linnaeus | |authority=Linnaeus | ||
| + | |rank=species | ||
| + | |publication_title=Sp. Pl. | ||
| + | |publication_place=2: 1093. 1753 | ||
}} | }} | ||
|synonyms={{Treatment/ID/Synonym | |synonyms={{Treatment/ID/Synonym | ||
|name=Dryopteris linnaeana | |name=Dryopteris linnaeana | ||
|authority=C. Christensen | |authority=C. Christensen | ||
| − | }}{{Treatment/ID/Synonym | + | |rank=species |
| + | }} {{Treatment/ID/Synonym | ||
|name=Lastrea dryopteris | |name=Lastrea dryopteris | ||
|authority=(Linnaeus) Bory | |authority=(Linnaeus) Bory | ||
| − | }}{{Treatment/ID/Synonym | + | |rank=species |
| + | }} {{Treatment/ID/Synonym | ||
|name=Phegopteris dryopteris | |name=Phegopteris dryopteris | ||
|authority=(Linnaeus) Fée | |authority=(Linnaeus) Fée | ||
| − | }}{{Treatment/ID/Synonym | + | |rank=species |
| + | }} {{Treatment/ID/Synonym | ||
|name=Thelypteris dryopteris | |name=Thelypteris dryopteris | ||
|authority=(Linnaeus) Slosson | |authority=(Linnaeus) Slosson | ||
| + | |rank=species | ||
}} | }} | ||
|hierarchy=Dryopteridaceae;Gymnocarpium;Gymnocarpium dryopteris | |hierarchy=Dryopteridaceae;Gymnocarpium;Gymnocarpium dryopteris | ||
| Line 32: | Line 39: | ||
}}<!-- | }}<!-- | ||
| − | --><span class="statement" id="st- | + | --><span class="statement" id="st-undefined" data-properties=""><b>Stems </b>0.5–1.5 mm diam.; scales 1–4 mm. <b>Fertile</b> leaves usually 12–42 cm. <b>Petiole</b> 9–28 cm, with sparse glandular hairs distally; scales 2–6 mm. <b>Blade</b> broadly deltate, 2-pinnate-pinnatifid, 3–14 cm, lax and delicate, abaxial surface and rachis glabrous or with sparse glandular hairs, adaxial surface glabrous. <b>Pinna</b> apex entire, rounded. <b>Proximal</b> pinnae 2–12 cm, ± perpendicular to rachis, with basiscopic pinnules ± perpendicular to costa; basal basiscopic pinnule usually sessile, pinnatifid or rarely pinnate-pinnatifid, if sessile then with basal basiscopic pinnulet often equaling or longer than adjacent pinnulet; 2d basal basiscopic pinnule sessile, with basal basiscopic pinnulet equaling or longer than adjacent pinnulet; basal acroscopic pinnule sessile, with basal basiscopic pinnulet longer than or equaling adjacent pinnulet. <b>Pinnae</b> of 2d pair usually sessile with basal basiscopic pinnule longer than or equaling adjacent pinnule and about equal to basal acroscopic pinnule; basal acroscopic pinnule equaling or slightly shorter than adjacent pinnule, often with entire, rounded apex. <b>Pinnae</b> of 3d pair sessile with basal basiscopic pinnule equaling adjacent pinnule and equaling basal acroscopic pinnules; basal acroscopic pinnule equaling or slightly shorter than adjacent pinnule. <b>Ultimate</b> segments of proximal pinnae oblong, entire to crenate, apex entire, rounded. <b>Spores</b> 34–39 µm. <b>2n</b> = 160.</span><!-- |
-->{{Treatment/Body | -->{{Treatment/Body | ||
|habitat=Cool, coniferous and mixed woods and at base of shale talus slopes | |habitat=Cool, coniferous and mixed woods and at base of shale talus slopes | ||
|elevation=0–3000 m | |elevation=0–3000 m | ||
| − | |distribution=Greenland;St. Pierre and Miquelon;Alta.;B.C.;Man.;N.B.;Nfld.;N.W.T.;N.S.;Ont.;P.E.I.;Que.;Sask.;Yukon;Alaska;Ariz.;Colo.;Conn.;Idaho;Iowa;Maine;Mass.;Mich.;Minn.;Mont.;N.H.;N.J.;N.Mex.;N.Y.;Ohio;Oreg.;Pa.;R.I.;S.Dak.;Vt.;Wash.;W.Va.;Wis.;Wyo.;n;c Europe;n Asia to China;Japan. | + | |distribution=Greenland;St. Pierre and Miquelon;Alta.;B.C.;Man.;N.B.;Nfld. and Labr.;N.W.T.;N.S.;Ont.;P.E.I.;Que.;Sask.;Yukon;Alaska;Ariz.;Colo.;Conn.;Idaho;Iowa;Maine;Mass.;Mich.;Minn.;Mont.;N.H.;N.J.;N.Mex.;N.Y.;Ohio;Oreg.;Pa.;R.I.;S.Dak.;Vt.;Wash.;W.Va.;Wis.;Wyo.;n;c Europe;n Asia to China;Japan. |
| − | |discussion=<p>Gymnocarpium dryopteris is a fertile allotetraploid species that arose following hybridization between G. appalachianum and G. disjunctum (see reticulogram). Its wide distribution over much of the north temperate zone has provided ample opportunity for secondary contact between G. dryopteris and each of its diploid parents, thereby resulting in a wide-ranging composite of abortive-spored triploid crosses (G. disjunctum × G. dryopteris and G. appalachianum × G. dryopteris). These relationships are shown on the diagram. Sterile triploid plants are not restricted only to areas where the range of the tetraploid overlaps with that of either diploid. Their broad distribution could be explained in part by their spores, which are of two types: malformed, black, and with very exaggerated perispores, or round with extensive netted perispores (K. M. Pryer and D. M. Britton 1983). The latter spore type is capable of germination and presumably permits the plants to reproduce apogamously. The name G. × brittonianum (Sarvela) Pryer & Haufler has been applied to the G. disjunctum × G. dryopteris hybrid formula (K. M. Pryer and C. H. Haufler 1993). The type of G. × brittonianum has aborted and round spores, and leaves that strongly resemble those of G. disjunctum. They are large, 3-pinnate-pinnatifid, and the second and third pairs of pinnae are sessile with basal basiscopic pinnules markedly longer than the basal acroscopic pinnules. Sterile triploid plants with a morphology similar to the type of G. × brittonianum are frequent. The biology of both of these cryptic hybrid taxa needs further study, which should lead to detailed morphologic descriptions and distribution maps.</p><!-- | + | |discussion=<p><i>Gymnocarpium dryopteris</i> is a fertile allotetraploid species that arose following hybridization between <i>G. appalachianum</i> and <i>G. disjunctum</i> (see reticulogram). Its wide distribution over much of the north temperate zone has provided ample opportunity for secondary contact between <i>G. dryopteris</i> and each of its diploid parents, thereby resulting in a wide-ranging composite of abortive-spored triploid crosses (<i>G. disjunctum</i> × <i>G. dryopteris</i> and <i>G. appalachianum</i> × <i>G. dryopteris</i>). These relationships are shown on the diagram. Sterile triploid plants are not restricted only to areas where the range of the tetraploid overlaps with that of either diploid. Their broad distribution could be explained in part by their spores, which are of two types: malformed, black, and with very exaggerated perispores, or round with extensive netted perispores (K. M. Pryer and D. M. Britton 1983). The latter spore type is capable of germination and presumably permits the plants to reproduce apogamously. The name G. × brittonianum (Sarvela) Pryer & Haufler has been applied to the <i>G. disjunctum</i> × <i>G. dryopteris</i> hybrid formula (K. M. Pryer and C. H. Haufler 1993). The type of G. × brittonianum has aborted and round spores, and leaves that strongly resemble those of <i>G. disjunctum</i>. They are large, 3-pinnate-pinnatifid, and the second and third pairs of pinnae are sessile with basal basiscopic pinnules markedly longer than the basal acroscopic pinnules. Sterile triploid plants with a morphology similar to the type of G. × brittonianum are frequent. The biology of both of these cryptic hybrid taxa needs further study, which should lead to detailed morphologic descriptions and distribution maps.</p><!-- |
| − | --><p>Gymnocarpium dryopteris also hybridizes with both G. jessoense subsp. parvulum and G. robertianum.</p> | + | --><p><i>Gymnocarpium dryopteris</i> also hybridizes with both <i>G. jessoense </i>subsp.<i> parvulum</i> and <i>G. robertianum</i>.</p> |
|tables= | |tables= | ||
|references= | |references= | ||
| Line 48: | Line 55: | ||
-->{{#Taxon: | -->{{#Taxon: | ||
name=Gymnocarpium dryopteris | name=Gymnocarpium dryopteris | ||
| − | |||
|authority=(Linnaeus) Newman | |authority=(Linnaeus) Newman | ||
|rank=species | |rank=species | ||
| Line 57: | Line 63: | ||
|habitat=Cool, coniferous and mixed woods and at base of shale talus slopes | |habitat=Cool, coniferous and mixed woods and at base of shale talus slopes | ||
|elevation=0–3000 m | |elevation=0–3000 m | ||
| − | |distribution=Greenland;St. Pierre and Miquelon;Alta.;B.C.;Man.;N.B.;Nfld.;N.W.T.;N.S.;Ont.;P.E.I.;Que.;Sask.;Yukon;Alaska;Ariz.;Colo.;Conn.;Idaho;Iowa;Maine;Mass.;Mich.;Minn.;Mont.;N.H.;N.J.;N.Mex.;N.Y.;Ohio;Oreg.;Pa.;R.I.;S.Dak.;Vt.;Wash.;W.Va.;Wis.;Wyo.;n;c Europe;n Asia to China;Japan. | + | |distribution=Greenland;St. Pierre and Miquelon;Alta.;B.C.;Man.;N.B.;Nfld. and Labr.;N.W.T.;N.S.;Ont.;P.E.I.;Que.;Sask.;Yukon;Alaska;Ariz.;Colo.;Conn.;Idaho;Iowa;Maine;Mass.;Mich.;Minn.;Mont.;N.H.;N.J.;N.Mex.;N.Y.;Ohio;Oreg.;Pa.;R.I.;S.Dak.;Vt.;Wash.;W.Va.;Wis.;Wyo.;n;c Europe;n Asia to China;Japan. |
|reference=None | |reference=None | ||
|publication title=Phytologist | |publication title=Phytologist | ||
|publication year=1851 | |publication year=1851 | ||
|special status= | |special status= | ||
| − | |source xml=https:// | + | |source xml=https://bitbucket.org/aafc-mbb/fna-data-curation/src/2e0870ddd59836b60bcf96646a41e87ea5a5943a/coarse_grained_fna_xml/V2/V2_786.xml |
|genus=Gymnocarpium | |genus=Gymnocarpium | ||
|species=Gymnocarpium dryopteris | |species=Gymnocarpium dryopteris | ||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
| − | |||
}}<!-- | }}<!-- | ||
-->[[Category:Treatment]][[Category:Gymnocarpium]] | -->[[Category:Treatment]][[Category:Gymnocarpium]] | ||
Latest revision as of 22:27, 20 February 2024
Stems 0.5–1.5 mm diam.; scales 1–4 mm. Fertile leaves usually 12–42 cm. Petiole 9–28 cm, with sparse glandular hairs distally; scales 2–6 mm. Blade broadly deltate, 2-pinnate-pinnatifid, 3–14 cm, lax and delicate, abaxial surface and rachis glabrous or with sparse glandular hairs, adaxial surface glabrous. Pinna apex entire, rounded. Proximal pinnae 2–12 cm, ± perpendicular to rachis, with basiscopic pinnules ± perpendicular to costa; basal basiscopic pinnule usually sessile, pinnatifid or rarely pinnate-pinnatifid, if sessile then with basal basiscopic pinnulet often equaling or longer than adjacent pinnulet; 2d basal basiscopic pinnule sessile, with basal basiscopic pinnulet equaling or longer than adjacent pinnulet; basal acroscopic pinnule sessile, with basal basiscopic pinnulet longer than or equaling adjacent pinnulet. Pinnae of 2d pair usually sessile with basal basiscopic pinnule longer than or equaling adjacent pinnule and about equal to basal acroscopic pinnule; basal acroscopic pinnule equaling or slightly shorter than adjacent pinnule, often with entire, rounded apex. Pinnae of 3d pair sessile with basal basiscopic pinnule equaling adjacent pinnule and equaling basal acroscopic pinnules; basal acroscopic pinnule equaling or slightly shorter than adjacent pinnule. Ultimate segments of proximal pinnae oblong, entire to crenate, apex entire, rounded. Spores 34–39 µm. 2n = 160.
Habitat: Cool, coniferous and mixed woods and at base of shale talus slopes
Elevation: 0–3000 m
Distribution
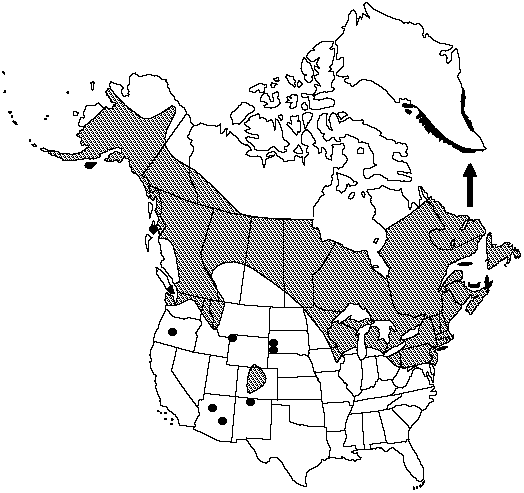
Greenland, St. Pierre and Miquelon, Alta., B.C., Man., N.B., Nfld. and Labr., N.W.T., N.S., Ont., P.E.I., Que., Sask., Yukon, Alaska, Ariz., Colo., Conn., Idaho, Iowa, Maine, Mass., Mich., Minn., Mont., N.H., N.J., N.Mex., N.Y., Ohio, Oreg., Pa., R.I., S.Dak., Vt., Wash., W.Va., Wis., Wyo., n, c Europe, n Asia to China, Japan.
Discussion
Gymnocarpium dryopteris is a fertile allotetraploid species that arose following hybridization between G. appalachianum and G. disjunctum (see reticulogram). Its wide distribution over much of the north temperate zone has provided ample opportunity for secondary contact between G. dryopteris and each of its diploid parents, thereby resulting in a wide-ranging composite of abortive-spored triploid crosses (G. disjunctum × G. dryopteris and G. appalachianum × G. dryopteris). These relationships are shown on the diagram. Sterile triploid plants are not restricted only to areas where the range of the tetraploid overlaps with that of either diploid. Their broad distribution could be explained in part by their spores, which are of two types: malformed, black, and with very exaggerated perispores, or round with extensive netted perispores (K. M. Pryer and D. M. Britton 1983). The latter spore type is capable of germination and presumably permits the plants to reproduce apogamously. The name G. × brittonianum (Sarvela) Pryer & Haufler has been applied to the G. disjunctum × G. dryopteris hybrid formula (K. M. Pryer and C. H. Haufler 1993). The type of G. × brittonianum has aborted and round spores, and leaves that strongly resemble those of G. disjunctum. They are large, 3-pinnate-pinnatifid, and the second and third pairs of pinnae are sessile with basal basiscopic pinnules markedly longer than the basal acroscopic pinnules. Sterile triploid plants with a morphology similar to the type of G. × brittonianum are frequent. The biology of both of these cryptic hybrid taxa needs further study, which should lead to detailed morphologic descriptions and distribution maps.
Gymnocarpium dryopteris also hybridizes with both G. jessoense subsp. parvulum and G. robertianum.
Selected References
None.