Bryoxiphium norvegicum
J. Linn. Soc., Bot. 12: 580. 1869,.

Plants 4–30 × 0.5–1.5 mm (not including flaring distal subulate leaves). Leaves somewhat scale-like proximally; median leaves oblong-lanceolate, rounded or nearly so, 1–2 mm, becoming apiculate to aristate; distal and gametoecial leaves 3–6 mm, long-subulate with ± twisted, ± hyaline, flexuous, smooth to spinose subula; costa ending few cells below apex in proximal and median leaves, ending in subula of distal and gametoecial leaves, interior cells of homogeneous stereids, exterior layer thinner-walled; abaxial lamella 1–6 cells high, sometimes absent, usually developed best on subulate leaves, usually ending well above insertion; interior laminal cells 11–40 × 9–18 µm; marginal cells 14–60 × 3.5–7.0 µm. Seta ca. 2 mm. Capsule ca. 1 mm. Spores 19–23 µm, slightly papillose.
Phenology: Capsules mature Jul.
Habitat: Usually on undersides of moist, shaded, sandstone ledges and cliffs, these occasionally calcareous and often overhanging streams, infrequently on bluffs and boulders of conglomerate, gneiss and quartzite, soil, and overturned tree bases
Elevation: low to high elevations (50-2800 m)
Distribution
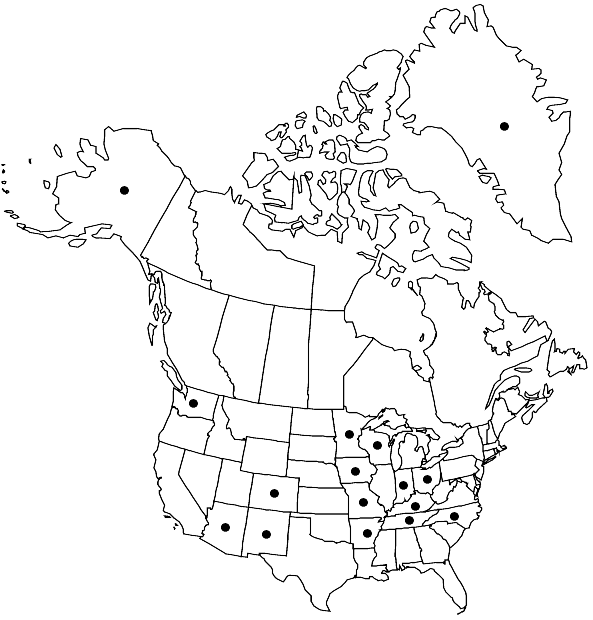
Greenland, Alaska, Ariz., Ark., Colo., Ind., Iowa, Ky., Minn., Mo., N.Mex., N.C., Ohio, Tenn., Wash., Wis., Mexico, West Indies (Dominican Republic), Asia, Atlantic Islands (Iceland, Madeira).
Discussion
Mature spores have been found only once in North America, in a sporophyte collected in the Wisconsin Dells in July. According to S. M. Hague and W. H. Welch (1951), antheridia are present from March to August, and archegonia are evident from May to August. In spite of the specific epithet, the original collection of this distinctive moss was made in Iceland. Usually bright green, Bryoxiphium norvegicum has on occasion been mistaken for grass seedlings. Bryoxiphium can also be confused with another moss genus, Fissidens. In both genera the leaves are distichously arranged, and the conduplicate (folded lengthwise) portion and weak abaxial lamella of the larger and better developed leaves of Bryoxiphium can be mistaken for the vaginant and abaxial laminae, respectively, of Fissidens. Costal structure of the two genera, however, is quite different, and species of Fissidens, with few exceptions, have a well-developed haplolepidous peristome.
Á. Löve and D. Löve (1953) recognized two subspecies of Bryoxiphium norvegicum, subsp. norvegicum and subsp. japonicum, distinguished by the degree of serrulation on the distal parts of the perichaetial leaves. Within subsp. norvegicum they recognized two varieties, norvegicum and mexicanum (the latter variety recognized at the species level by H. A. Crum in A. J. Sharp et al. (1994), based on differences in the length of marginal cells in perichaetial leaves. According to Löve and Löve, North American populations north of Mexico belong to var. norvegicum, having perichaetial leaves only slightly serrulate and marginal cells much longer than the interior laminal cells. A report of B. norvegicum from Pennsylvania cannot be substantiated.
Selected References
None.