Difference between revisions of "Weissia phascopsis"
Monogr. Syst. Bot. Missouri Bot. Gard. 11: 196. 1985,.
imported>Volume Importer |
imported>Volume Importer |
||
| Line 63: | Line 63: | ||
|publication year= | |publication year= | ||
|special status= | |special status= | ||
| − | |source xml=https:// | + | |source xml=https://bitbucket.org/aafc-mbb/fna-data-curation/src/2e0870ddd59836b60bcf96646a41e87ea5a5943a/coarse_grained_fna_xml/V27/V27_741.xml |
|subfamily=Pottiaceae subfam. Trichostomoideae | |subfamily=Pottiaceae subfam. Trichostomoideae | ||
|genus=Weissia | |genus=Weissia | ||
Revision as of 21:28, 5 November 2020
Leaves long-lanceolate, base broadly elliptical, shoulders absent, distal laminal margins strongly and sharply incurved or inrolled, apex narrowly channeled, acute, mucro strong, of 6–10 cells; costal adaxial stereid band smaller than the abaxial; distal laminal cells 8–10 µm wide. Sexual condition cladautoicous. Seta short, 0.015–0.017 cm not including the vaginula. Capsule cleistocarpic, elliptic to ovoid with a long-rostrate apiculus.
Phenology: Capsules mature winter–spring.
Habitat: Soil
Elevation: moderate elevations
Distribution
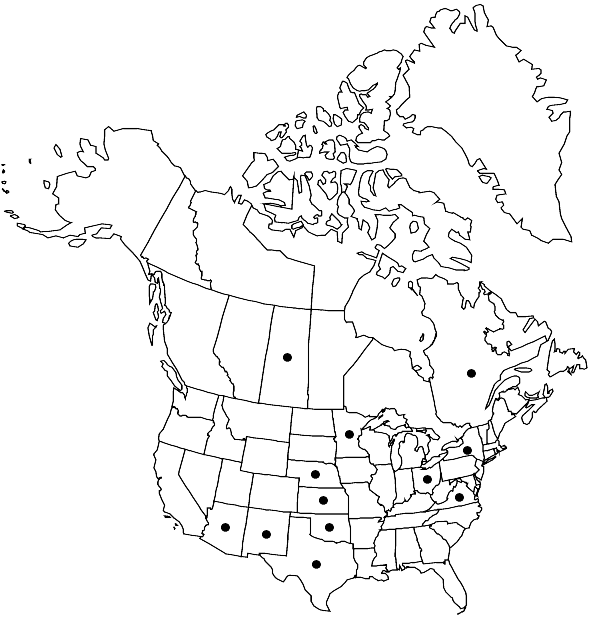
Que., Sask., Ariz., Kans., Minn., Nebr., N.Mex., N.Y., Ohio, Okla., Tex., Va., Europe.
Discussion
Some capsules of Weissia phascopsis show rudimentary peristomes, of 1–8 teeth, each of 1–2 articulations, which may vary in a tuft, and aborted spores were not seen. The possible hybrid origin of this species has been discussed by A. L. Andrews (1920, 1922, 1924, 1933). C. Williams (1966) found possible natural hybrids in Ontario between W. controversa and W. muhlenbergiana, which differed from W. phascopsis by the large number of abortive spores and rudimentary peristome teeth (16 with 4 articulations). The difference in number of teeth and articulations indicates that W. phascopsis is either not a hybrid or it has backcrossed to W. muhlenbergiana. H. A. Crum and L. E. Anderson (1981) gave a clear summary of the problem. Species previously placed in Astomum are here presented as separate species pending a better analysis of the genetic basis of differences in traits. The gametophyte of W. phascopsis is much the same as that of W. controversa, which supports a theory of hybrid origin of the sporophyte. Further evidence is the occasional rudimentary peristome. The sexuality, however, is cladautoicous. This, and the multiple identical sporophytes per turf, each with no or few aborted spores, argue against outcrossing. There is no evidence for differential timing of maturation and dehiscence of archegonia and antheridia.
Selected References
None.