Difference between revisions of "Draba lactea"
Mém. Soc. Imp. Naturalistes Moscou 5: 104. 1817.
FNA>Volume Importer |
FNA>Volume Importer |
||
| Line 11: | Line 11: | ||
|name=Draba allenii | |name=Draba allenii | ||
|authority=Fernald | |authority=Fernald | ||
| − | }}{{Treatment/ID/Synonym | + | }} {{Treatment/ID/Synonym |
|name=Draba boecheri | |name=Draba boecheri | ||
|authority=Gjaerevoll & Ryvarden | |authority=Gjaerevoll & Ryvarden | ||
| − | }}{{Treatment/ID/Synonym | + | }} {{Treatment/ID/Synonym |
|name=Draba fernaldiana | |name=Draba fernaldiana | ||
|authority=Polunin | |authority=Polunin | ||
| Line 32: | Line 32: | ||
|elevation=0-2000 m | |elevation=0-2000 m | ||
|distribution=Greenland;Nfld. and Labr.;N.W.T.;Nunavut;Que.;Yukon;Alaska;Europe (n Finland;Norway;w Sweden);Asia (Russian Far East;c;n Siberia);Atlantic Islands (Iceland);circumpolar. | |distribution=Greenland;Nfld. and Labr.;N.W.T.;Nunavut;Que.;Yukon;Alaska;Europe (n Finland;Norway;w Sweden);Asia (Russian Far East;c;n Siberia);Atlantic Islands (Iceland);circumpolar. | ||
| − | |discussion=<p>T. W. Böcher (1966) postulated that Draba lactea originated from hybridization between D. fladnizensis and D. nivalis, but A.-C. Scheen et al. (2002) showed that it is more closely allied to D. subcapitata. By contrast, H. H. Grundt et al. (2004) concluded that hexaploid D. lactea originated from tetraploids of the same species, which in turn originated from the diploid D. palanderiana lineage. They suggested that D. lactea probably originated multiple times in the Beringian area and migrated to reach its present circumpolar distribution. The hexaploids are distributed throughout the species range, whereas the tetraploids are known only from Alaska and the Russian Far East (Grundt et al. 2005b).</p><!-- | + | |discussion=<p>T. W. Böcher (1966) postulated that <i>Draba lactea</i> originated from hybridization between <i>D. fladnizensis</i> and <i>D. nivalis</i>, but A.-C. Scheen et al. (2002) showed that it is more closely allied to <i>D. subcapitata</i>. By contrast, H. H. Grundt et al. (2004) concluded that hexaploid <i>D. lactea</i> originated from tetraploids of the same species, which in turn originated from the diploid <i>D. palanderiana</i> lineage. They suggested that <i>D. lactea</i> probably originated multiple times in the Beringian area and migrated to reach its present circumpolar distribution. The hexaploids are distributed throughout the species range, whereas the tetraploids are known only from Alaska and the Russian Far East (Grundt et al. 2005b).</p><!-- |
| − | --><p>Draba fernaldiana, which was collected from Southampton Island (Nunavut), was not mentioned by R. C. Rollins (1993). The plants are completely glabrous except for leaf margins, which are ciliate with simple and sparse 2-rayed trichomes. The taxon resembles some forms of D. lactea and is tentatively herein included within that species. The only conflict in such placement is petal color, which was listed in the original description of D. fernaldiana as pale yellow instead of white.</p><!-- | + | --><p><i>Draba</i> fernaldiana, which was collected from Southampton Island (Nunavut), was not mentioned by R. C. Rollins (1993). The plants are completely glabrous except for leaf margins, which are ciliate with simple and sparse 2-rayed trichomes. The taxon resembles some forms of <i>D. lactea</i> and is tentatively herein included within that species. The only conflict in such placement is petal color, which was listed in the original description of D. fernaldiana as pale yellow instead of white.</p><!-- |
| − | --><p>Glabrous or glabrescent forms of Draba lactea are quite common in the Canadian Arctic Archipelago, whereas pubescent forms predominate in Alaska and the Russian Far East.</p> | + | --><p>Glabrous or glabrescent forms of <i>Draba lactea</i> are quite common in the Canadian Arctic Archipelago, whereas pubescent forms predominate in Alaska and the Russian Far East.</p> |
|tables= | |tables= | ||
|references= | |references= | ||
| Line 58: | Line 58: | ||
|publication year=1817 | |publication year=1817 | ||
|special status= | |special status= | ||
| − | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/ | + | |source xml=https://jpend@bitbucket.org/aafc-mbb/fna-data-curation.git/src/8f726806613d60c220dc4493de13607dd3150896/coarse_grained_fna_xml/V7/V7_421.xml |
|tribe=Brassicaceae tribe Arabideae | |tribe=Brassicaceae tribe Arabideae | ||
|genus=Draba | |genus=Draba | ||
Revision as of 17:57, 18 September 2019
Perennials; (cespitose); caudex branched (with persistent petiole remains, branches sometimes terminating in sterile rosettes); scapose. Stems unbranched, 0.2–1.1(–1.5) dm,glabrous throughout or sparsely pubescent proximally, trichomes short-stalked, substellate, 2–8-rayed, (non-crisped), 0.5–0.3 mm. Basal leaves rosulate; petiolate; petiole (persistent, strongly thickened), margin usually ciliate, (trichomes usually simple and 2-rayed, 0.3–1 mm); blade oblanceolate to obovate, (0.3–)0.5–1.1(–1.7) cm × (1–)2–6 mm, margins usually entire, rarely denticulate, (sometimes ciliate), surfaces sometimes pubescent with stellate to subdendritic, 4–12-rayed, (non-crisped) trichomes, 0.1–0.4 mm, (midvein persistent, prominent, strongly thickened). Cauline leaves 0 (or, rarely, 1 as a bract). Racemes 2–8(–12)-flowered, ebracteate, elongated in fruit; rachis not flexuous, usually glabrous, rarely sparsely pubescent as stem basally. Fruiting pedicels divaricate-ascending, straight, (1–)2–5(–10) mm, glabrous. Flowers: sepals ovate, 1.8–3 mm, usually glabrous, rarely sparsely pubescent subapically, (trichomes simple); petals white, obovate, 3–5 × 1.8–3 mm; anthers ovate, 0.3–0.4 mm. Fruits oblong to elliptic-lanceolate or ovate to broadly so, plane, flattened, 4–8 × (1.5–)2–3 mm; valves glabrous; ovules (10–)14–22(–26) per ovary; style 0.1–0.4 mm. Seeds ovoid, 0.8–1.1 × 0.5–0.6 mm. 2n = 32, 48.
Phenology: Flowering Jun–Aug.
Habitat: Rock outcrops, talus, rocky hillsides and ridges, open gravelly areas, seepage swales, meadows
Elevation: 0-2000 m
Distribution
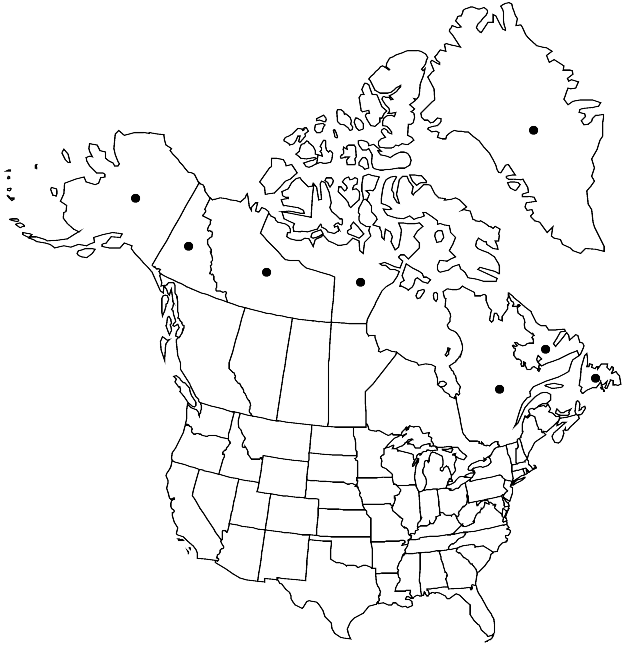
Greenland, Nfld. and Labr., N.W.T., Nunavut, Que., Yukon, Alaska, Europe (n Finland, Norway, w Sweden), Asia (Russian Far East, c, n Siberia), Atlantic Islands (Iceland), circumpolar.
Discussion
T. W. Böcher (1966) postulated that Draba lactea originated from hybridization between D. fladnizensis and D. nivalis, but A.-C. Scheen et al. (2002) showed that it is more closely allied to D. subcapitata. By contrast, H. H. Grundt et al. (2004) concluded that hexaploid D. lactea originated from tetraploids of the same species, which in turn originated from the diploid D. palanderiana lineage. They suggested that D. lactea probably originated multiple times in the Beringian area and migrated to reach its present circumpolar distribution. The hexaploids are distributed throughout the species range, whereas the tetraploids are known only from Alaska and the Russian Far East (Grundt et al. 2005b).
Draba fernaldiana, which was collected from Southampton Island (Nunavut), was not mentioned by R. C. Rollins (1993). The plants are completely glabrous except for leaf margins, which are ciliate with simple and sparse 2-rayed trichomes. The taxon resembles some forms of D. lactea and is tentatively herein included within that species. The only conflict in such placement is petal color, which was listed in the original description of D. fernaldiana as pale yellow instead of white.
Glabrous or glabrescent forms of Draba lactea are quite common in the Canadian Arctic Archipelago, whereas pubescent forms predominate in Alaska and the Russian Far East.
Selected References
None.