Difference between revisions of "Cirsium scariosum var. scariosum"
FNA>Volume Importer |
imported>Volume Importer |
||
| (6 intermediate revisions by 2 users not shown) | |||
| Line 1: | Line 1: | ||
{{Treatment/ID | {{Treatment/ID | ||
|accepted_name=Cirsium scariosum var. scariosum | |accepted_name=Cirsium scariosum var. scariosum | ||
| − | |accepted_authority= | + | |accepted_authority= |
|publications= | |publications= | ||
| + | |special_status={{Treatment/ID/Special_status | ||
| + | |code=F | ||
| + | |label=Illustrated | ||
| + | }}{{Treatment/ID/Special_status | ||
| + | |code=E | ||
| + | |label=Endemic | ||
| + | }} | ||
|basionyms= | |basionyms= | ||
|synonyms={{Treatment/ID/Synonym | |synonyms={{Treatment/ID/Synonym | ||
|name=Cirsium butleri | |name=Cirsium butleri | ||
|authority=(Rydberg) Petrak | |authority=(Rydberg) Petrak | ||
| − | }}{{Treatment/ID/Synonym | + | |rank=species |
| + | }} {{Treatment/ID/Synonym | ||
|name=Cirsium lacerum | |name=Cirsium lacerum | ||
|authority=(Rydberg) Petrak | |authority=(Rydberg) Petrak | ||
| − | }}{{Treatment/ID/Synonym | + | |rank=species |
| + | }} {{Treatment/ID/Synonym | ||
|name=Cirsium magnificum | |name=Cirsium magnificum | ||
|authority=(A. Nelson) Petrak | |authority=(A. Nelson) Petrak | ||
| − | }}{{Treatment/ID/Synonym | + | |rank=species |
| + | }} {{Treatment/ID/Synonym | ||
|name=Cirsium minganense | |name=Cirsium minganense | ||
|authority=Victorin | |authority=Victorin | ||
| + | |rank=species | ||
}} | }} | ||
|hierarchy=Asteraceae;Asteraceae tribe Cardueae;Cirsium;Cirsium scariosum;Cirsium scariosum var. scariosum | |hierarchy=Asteraceae;Asteraceae tribe Cardueae;Cirsium;Cirsium scariosum;Cirsium scariosum var. scariosum | ||
| Line 24: | Line 35: | ||
}}<!-- | }}<!-- | ||
| − | --><span class="statement" id="st-undefined" data-properties=""><b>Plants </b>erect, caulescent or occasionally acaulescent, 15–200 cm. <b>Stems</b> usually simple, very leafy, glabrous or villous to tomentose with septate trichomes. <b>Leaves</b>: blades linear to oblong, oblanceolate, or narrowly elliptic, pinnately lobed or often unlobed, larger leaf spines usually 1 cm or less, abaxial faces glabrous to gray-tomentose, adaxial glabrous or villous with septate trichomes; distal narrow, ± thin, often unpigmented proximally or tinged pink or purplish, spines numerous, weak. <b>Heads</b> 1–10+, sessile or short-pedunculate, tightly clustered at stem tips, usually subtended and ± overtopped by crowded, distal leaves. <b>Involucres</b> 2–3.5 cm. <b>Phyllaries</b>: outer and mid lanceolate to ovate, spines slender to stout, 1–8 mm; apices of inner acuminate and entire or abruptly expanded into scarious, erose-toothed appendages. <b>Corollas</b> white to purple, 20–28 mm, tubes 9–14.5 mm, throats 6–10 mm, lobes 3.5–6 mm; style tips 3–6.5 mm. <b>Cypselae</b> 4.5–6.5 mm; pappi 17–25 mm. <b>2n</b> = 34 (as C. foliosum), 36.</span><!-- | + | --><span class="statement" id="st-undefined" data-properties=""><b>Plants </b>erect, caulescent or occasionally acaulescent, 15–200 cm. <b>Stems</b> usually simple, very leafy, glabrous or villous to tomentose with septate trichomes. <b>Leaves</b>: blades linear to oblong, oblanceolate, or narrowly elliptic, pinnately lobed or often unlobed, larger leaf spines usually 1 cm or less, abaxial faces glabrous to gray-tomentose, adaxial glabrous or villous with septate trichomes; distal narrow, ± thin, often unpigmented proximally or tinged pink or purplish, spines numerous, weak. <b>Heads</b> 1–10+, sessile or short-pedunculate, tightly clustered at stem tips, usually subtended and ± overtopped by crowded, distal leaves. <b>Involucres</b> 2–3.5 cm. <b>Phyllaries</b>: outer and mid lanceolate to ovate, spines slender to stout, 1–8 mm; apices of inner acuminate and entire or abruptly expanded into scarious, erose-toothed appendages. <b>Corollas</b> white to purple, 20–28 mm, tubes 9–14.5 mm, throats 6–10 mm, lobes 3.5–6 mm; style tips 3–6.5 mm. <b>Cypselae</b> 4.5–6.5 mm; pappi 17–25 mm. <b>2n</b> = 34 (as <i>C. foliosum</i>), 36.</span><!-- |
-->{{Treatment/Body | -->{{Treatment/Body | ||
| Line 31: | Line 42: | ||
|elevation=0 (Quebec) or 600–2800 m | |elevation=0 (Quebec) or 600–2800 m | ||
|distribution=Alta.;B.C.;Que.;Calif.;Colo.;Idaho;Mont.;Oreg.;Utah;Wash.;Wyo. | |distribution=Alta.;B.C.;Que.;Calif.;Colo.;Idaho;Mont.;Oreg.;Utah;Wash.;Wyo. | ||
| − | |discussion=<p>The presence of Cirsium scariosum on the islands of the Mingan Archipelago in Quebec, some 3200 km east of the Rocky Mountains populations, has led to alternative hypotheses regarding the disjunction. Frère Marie-Victorin (1925) hypothesized that the disjunct distribution of C. minganense from what he called C. foliosum (Hooker) Candolle was a result of migration during deglaciation (18,000 to ca. 8000 BP) from a glacial refugium in western North America to eastern Canada in the barren habitats along the receding ice front. Later (1938) he presented a second hypothesis that Pleistocene glacial events had divided a preglacial range into vicariant populations that survived in separate refugia in western and eastern regions. R. J. Moore and C. Frankton (1967) argued that the disjunction is modern, resulting from a chance introduction of C. scariosum from western North America to Quebec in the early twentieth century. They reached this conclusion because early collectors that had visited the Mingan Archipelago had failed to collect this conspicuous thistle.</p><!-- | + | |discussion=<p>The presence of <i>Cirsium scariosum</i> on the islands of the Mingan Archipelago in Quebec, some 3200 km east of the Rocky Mountains populations, has led to alternative hypotheses regarding the disjunction. Frère Marie-Victorin (1925) hypothesized that the disjunct distribution of C. minganense from what he called <i>C. foliosum</i> (Hooker) Candolle was a result of migration during deglaciation (18,000 to ca. 8000 BP) from a glacial refugium in western North America to eastern Canada in the barren habitats along the receding ice front. Later (1938) he presented a second hypothesis that Pleistocene glacial events had divided a preglacial range into vicariant populations that survived in separate refugia in western and eastern regions. R. J. Moore and C. Frankton (1967) argued that the disjunction is modern, resulting from a chance introduction of <i>C. scariosum</i> from western North America to Quebec in the early twentieth century. They reached this conclusion because early collectors that had visited the Mingan Archipelago had failed to collect this conspicuous thistle.</p><!-- |
| − | --><p>Hybrids are known between Cirsium scariosum var. scariosum and C. eatonii var. murdockii in northern Wyoming.</p> | + | --><p>Hybrids are known between <i>Cirsium scariosum </i>var.<i> scariosum</i> and <i>C. eatonii </i>var.<i> murdockii</i> in northern Wyoming.</p> |
|tables= | |tables= | ||
|references= | |references= | ||
| Line 41: | Line 52: | ||
-->{{#Taxon: | -->{{#Taxon: | ||
name=Cirsium scariosum var. scariosum | name=Cirsium scariosum var. scariosum | ||
| − | + | |authority= | |
| − | |authority= | ||
|rank=variety | |rank=variety | ||
|parent rank=species | |parent rank=species | ||
| Line 55: | Line 65: | ||
|publication title= | |publication title= | ||
|publication year= | |publication year= | ||
| − | |special status= | + | |special status=Illustrated;Endemic |
| − | |source xml=https:// | + | |source xml=https://bitbucket.org/aafc-mbb/fna-data-curation/src/2e0870ddd59836b60bcf96646a41e87ea5a5943a/coarse_grained_fna_xml/V19-20-21/V19_151.xml |
|tribe=Asteraceae tribe Cardueae | |tribe=Asteraceae tribe Cardueae | ||
|genus=Cirsium | |genus=Cirsium | ||
Latest revision as of 19:48, 5 November 2020

Plants erect, caulescent or occasionally acaulescent, 15–200 cm. Stems usually simple, very leafy, glabrous or villous to tomentose with septate trichomes. Leaves: blades linear to oblong, oblanceolate, or narrowly elliptic, pinnately lobed or often unlobed, larger leaf spines usually 1 cm or less, abaxial faces glabrous to gray-tomentose, adaxial glabrous or villous with septate trichomes; distal narrow, ± thin, often unpigmented proximally or tinged pink or purplish, spines numerous, weak. Heads 1–10+, sessile or short-pedunculate, tightly clustered at stem tips, usually subtended and ± overtopped by crowded, distal leaves. Involucres 2–3.5 cm. Phyllaries: outer and mid lanceolate to ovate, spines slender to stout, 1–8 mm; apices of inner acuminate and entire or abruptly expanded into scarious, erose-toothed appendages. Corollas white to purple, 20–28 mm, tubes 9–14.5 mm, throats 6–10 mm, lobes 3.5–6 mm; style tips 3–6.5 mm. Cypselae 4.5–6.5 mm; pappi 17–25 mm. 2n = 34 (as C. foliosum), 36.
Phenology: Flowering summer (Jun–Sep).
Habitat: Moist, sometimes saline soils, meadows, ditches, stream banks, forest openings, sagebrush zone to subalpine forests
Elevation: 0 (Quebec) or 600–2800 m
Distribution
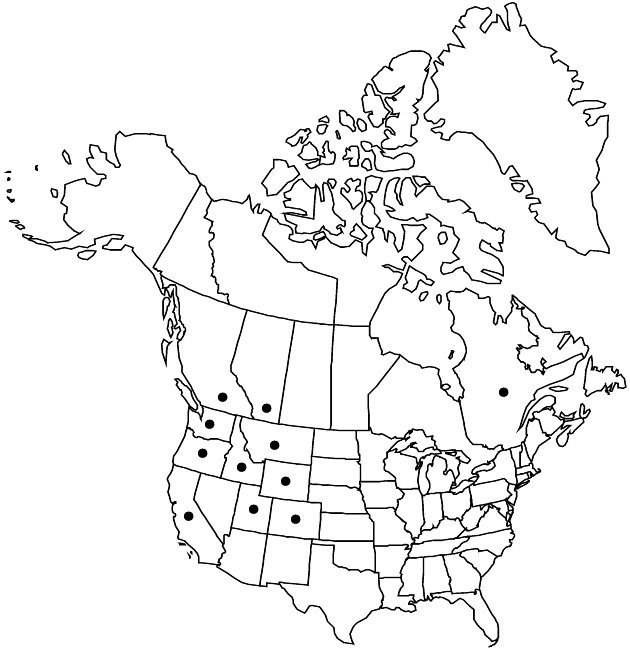
Alta., B.C., Que., Calif., Colo., Idaho, Mont., Oreg., Utah, Wash., Wyo.
Discussion
The presence of Cirsium scariosum on the islands of the Mingan Archipelago in Quebec, some 3200 km east of the Rocky Mountains populations, has led to alternative hypotheses regarding the disjunction. Frère Marie-Victorin (1925) hypothesized that the disjunct distribution of C. minganense from what he called C. foliosum (Hooker) Candolle was a result of migration during deglaciation (18,000 to ca. 8000 BP) from a glacial refugium in western North America to eastern Canada in the barren habitats along the receding ice front. Later (1938) he presented a second hypothesis that Pleistocene glacial events had divided a preglacial range into vicariant populations that survived in separate refugia in western and eastern regions. R. J. Moore and C. Frankton (1967) argued that the disjunction is modern, resulting from a chance introduction of C. scariosum from western North America to Quebec in the early twentieth century. They reached this conclusion because early collectors that had visited the Mingan Archipelago had failed to collect this conspicuous thistle.
Hybrids are known between Cirsium scariosum var. scariosum and C. eatonii var. murdockii in northern Wyoming.
Selected References
None.